


-
结直肠癌是一种常见的恶性肿瘤,其致死率在恶性肿瘤中占第3位,仅次于肺癌和乳腺癌[1]。目前,放疗在结直肠癌的治疗过程中发挥着不可替代的作用[2]。然而,结直肠癌对放疗的抗性严重地影响了放疗患者的治疗效果与生存质量,其放疗抗性机制尚不明确[3-4]。基因表达数据库(Gene Expression Omnibus,GEO)是当今较大、且全面的公共基因表达数据库,为基因研究提供了大量的高通量数据[5]。本研究中我们主要利用GEO数据库筛选结直肠癌辐射耐受细胞中的差异表达基因,从分子水平上初步探讨与结直肠癌抗辐射相关的潜在基因,为下一步的实验研究提供理论基础。
-
结直肠癌辐射耐受细胞的基因表达谱数据集GSE43206来源于GEO数据库( https://www.ncbi.nlmnih.gov/geo/. ),芯片数据采用的平台为GPL6244。该数据集由6个样本构成,包括照射前后的结直肠癌细胞,其中,GSM105861、GSM105863、GSM105865是未照射的结直肠癌细胞,而GSM105862、GSM105864、GSM105866是经辐射诱导的耐辐射结直肠癌细胞。照射条件是来自137Cs源的γ射线,照射总剂量为80 Gy。
-
首先,我们从GEO数据库下载原始的GSE43206基因芯片数据,根据平台GPL6244提供的注释文件,对基因功能进行注释,将探针名转化为对应的基因名,形成基因表达矩阵。随后,利用R语言及相关的Bioconductor软件包,对芯片数据进行预处理分析[6],经过数据过滤,log2转换,采用K近邻法补充缺失值[7-8],再采用RMA算法对数据进行归一化处理。利用R语言中的limma包[9]对标准化数据进行倍数差异分析和t检验分析,从标准化的数据中筛选出差异表达基因,筛选条件:P < 0.05和|log2 fold change|≥1。同时,为了确定样本中差异基因的筛选情况,利用pheatmap包对部分差异基因进行分层聚类。差异倍数(fold change)=辐射耐受株细胞中基因相对表达量/未照射细胞中基因相对表达量。
-
利用R语言中的clusterProfiler包和ggplot2包,对差异基因中的编码基因进行基因本体论(gene ontology,GO)功能富集分析和京都基因和基因组百科全书(kyoto encyclopedia of genes and genomes,KEGG)通路分析。其中P < 0.01和Q < 0.05作为筛选条件。随后利用公共人源蛋白相关数据库[10](https://www.hprd.org/),对差异基因中的编码基因构建蛋白相互作用(protein-protein interaction,PPI)网络,结果在cytoscape3.6.0软件[11-12]中实现可视化。最后,通过cytoscape 3.6.0软件的cytoHubba插件[13-14]中的点度心性[16]、中介中心性[17]等方法确定PPI网络中的关键基因。
-
将细胞接种于6孔板中,每组3个复孔。根据不同的处理方法,将人结肠癌HCT116细胞(购自美国ATCC细胞库)分为两组,照射组和未照射组。24 h后,采用137Cs γ射线照射源(购自加拿大原子能有限公司),照射剂量分别为0和5 Gy,吸收剂量率为0.99 Gy /min。收集照射后24 h的人结肠癌HCT116细胞,Trizol法提取细胞总RNA,用反转录试剂盒(购自日本TaKaRa公司)反转录mRNA生成cDNA,采用实时荧光定量PCR检测仪(购自杭州博日科技有限公司,型号:FQD-96A)检测照射后人结直肠癌HCT116细胞中NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR等基因的mRNA表达,选择GAPDH基因作为mRNA的内参基因。扩增条件如下,恒温段:50 ℃、2 min,95 ℃、10 min;循环段:95℃、15 s,60 ℃、1 min;熔解段:95 ℃、15 s,60 ℃、1 min,95 ℃、15 s,循环40次。数据采用2-ΔΔCt法进行分析。
GAPDH引物序列:
正向:5’-GGAGCGAGATCCCTCCAAAAT-3’
反向:5’-GGCTGTTGTCATACTTCTCATGC-3’
NDRG1引物序列:
正向:5’-CTCCTGCAAGAGTTTGATGTCC-3’
反向:5’-TCATGCCGATGTCATGGTAGG-3’
PAG1引物序列:
正向:5’-TTCAGCCGTTCAGTTACTAGCC-3’
反向:5’-TGGACTTCCTCGTAATGCTGC-3’
LRP1引物序列:
正向:5’-CTATCGACGCCCCTAAGACTT-3’
反向:5’-CATCGCTGGGCCTTACTCT-3’
PIM1引物序列:
正向:5’-GAGAAGGACCGGATTTCCGAC-3’
反向:5’-CAGTCCAGGAGCCTAATGACG-3’
LDLR引物序列:
正向:5’-ACGGCGTCTCTTCCTATGACA-3’
反向:5’-CCCTTGGTATCCGCAACAGA-3’
PLAUR引物序列:
正向:5’-TGTAAGACCAACGGGGATTGC-3’
反向:5’-AGCCAGTCCGATAGCTCAGG-3’
-
采用SPSS19.0软件中的Student t-test进行统计学分析。每组实验至少重复3次。原始数据均服从正态分布且方差齐,采用t检验比较组间差异,P<0.05表示差异有统计学意义。
-
经过数据归一化处理,每组样本数据的中位值保持一致(归一化后中位值为52.48),消除了因个体基因组间不同所造成的基因间的差别,显示出良好的标准化程度,使得后续数据分析能够获得真实的处理结果。
-
应用R语言中的limma包共筛选出差异基因101个,其中67个基因上调,占66.34%;34个基因下调,占33.66%。对前15个差异基因进行聚类分析结果见图 1所示,上调基因和下调基因中所对应的前15个差异基因可以被明显识别并加以区分。

图 1 差异基因筛选的聚类分析图
Figure 1. Screening differentially expressed genes
-
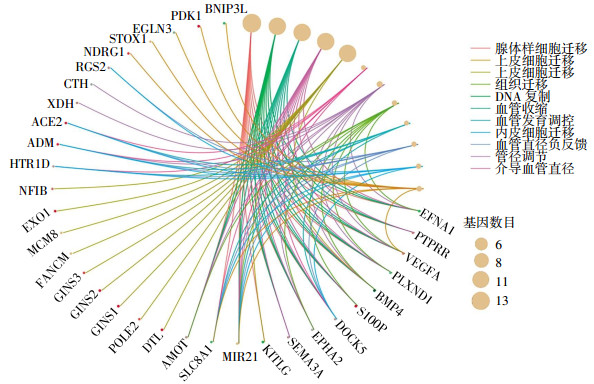
GO富集分析结果显示,差异基因富集在与细胞迁移、DNA复制、血管收缩等相关的生物学过程中,结果见图 2。KEGG通路分析显示,经过功能注释的差异基因主要富集在乏氧诱导因子1信号通路(VEGFA、EGLN3、HK2、ENO2、PDK1)、胆固醇代谢(SOAT1、LRP1、LDLR)等相关通路上。

图 2 基因本体论富集分析图
Figure 2. Gene Ontology enrichment analysis
-
结果显示,PPI网络主要由122个基因和123种相互作用构成。同时,为了描述网络中存在的重要节点,我们采用点度中心性[16]、中介中心性[17]等方法,筛选出排名前10的基因,结果见表 1。通过对得到的Top基因取交集,最终得到在PPI网络中发挥关键作用的6个关键基因为NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR基因。
基因名 点度中心性 基因名 点度中心性 NDRG1 18 NDRG1 29.78 PAG1 16 LDLR 29 ENO2 12 HSPA5 25.95 RGS2 10 LRP1 24.87 LRP1 9 PIM1 25.03 PIM1 8 HSP90AA1 23.4 VEGFA 8 PAG1 22.57 LDLR 7 PLAUR 22.45 PLAUR 7 EFNA1 22.33 EPHA2 6 EPHA3 22.28 表 1 蛋白相互作用网络中根据节点中心度筛选的前10个基因
Table 1. The top 10 genes in protein-protein interaction network Network topology parameters
-
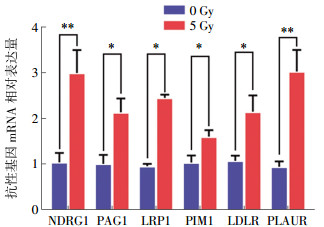
结果显示,在人结肠癌HCT16细胞中,NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR等潜在抗性基因接受照射后,mRNA表达量显著上升,与照射前相比,差异均有统计学意义(t=49.981,P < 0.01;t=26.420、28.698、21.358、23.545,均P < 0.05;t=50.601,P < 0.01)。虽然这些抗性基因在照射后的mRNA相对表达水平与生物信息学预测结果相一致,但仍略有差别。实时荧光定量PCR实验证实,NDRG1和PLAUR(t=49.981、50.601,均P < 0.01)的mRNA相对表达水平在照射后变化明显,约为3倍差异的表达量变化,而基因芯片结果显示的差异变化明显的抗性基因为NDRG1和PAG1,差异倍数分别为2.60倍和1.96倍。结果见图 3。

图 3 潜在辐射抗性基因mRNA相对表达量
Figure 3. The relative expression of potential radiation resistance genes
-
结直肠癌是常见的胃肠道肿瘤之一。尽管在筛查、诊断和治疗方面取得了进展,但其致死率在恶性肿瘤中仍占第3位[1]。目前,手术切除仍旧作为治疗结直肠癌的主要方法,同时与放疗、化疗及靶向治疗等联合应用可提高疗效和降低转移的可能性。如术前放疗可有效缩小肿块、提高保肛率,并控制癌细胞可能的转移,从而为后续的手术治疗提供有利条件。而在术后使用放疗可以降低局部复发率,延长生存期[2]。遗憾的是,部分结直肠癌患者在临床上表现为放疗不敏感甚至放疗抗拒,严重影响了后续的治疗效果。因此,对结直肠癌患者放疗敏感性的探索是近年来的研究热点。
本研究在分子水平上,利用生物信息学技术初步探索结直肠癌抗辐射细胞中的差异表达基因。首先,在公共数据库GEO中下载相关的芯片数据,通过对芯片数据的预处理并应用R语言中的limma包,共筛选出101个差异基因,其中包含67个上调的差异基因和34个下调的差异基因。对这些差异基因进行GO富集分析和KEGG通路分析,结果发现,这些差异基因富集在与细胞迁移、DNA复制、血管收缩等相关的生物学进程中以及乏氧诱导因子1信号通路、胆固醇代谢等相关通路上。多项研究表明,电离辐射可能通过微环境、细胞-细胞连接、细胞外基质连接、蛋白酶分泌和诱导上皮-间质转化(endothelial mesenchymal transdifferentiation,EMT)等复杂的途径促进肿瘤细胞的迁移和侵袭[19-20]。大量的化学物质和一些物理因素,如紫外线和γ射线,可影响DNA复制,诱导DNA发生突变,在人类癌症的发生发展中发挥着至关重要的作用[21]。另外,电离辐射通过诱导EMT,有助于癌细胞的转移和侵袭[20]。Lee等[22]证实,乏氧诱导因子1信号通路的激活在辐射诱导的EMT中起重要作用。
通过对差异基因构建PPI网络,我们发现NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR共6个基因在PPI网络中发挥着主导作用。研究发现,NDRG1基因是一种抑癌基因,可以抑制肿瘤转移、癌细胞的迁移和侵袭[23]。在p53介导的细胞凋亡中扮演着至关重要的作用。Ma等[24]研究证实,降低NDRG1的表达可以促进结肠癌细胞的EMT,通过激活核因子κB信号通路实现。目前关于NDRG1基因是否与放射敏感性相关的研究尚未见报道。PAG1基因主要编码一种与糖脂富集相关的磷酸化蛋白。在对神经母细胞瘤的研究中发现,通过降低PAG1的表达可以激活蛋白激酶B和酪氨酸蛋白激酶c-Src的表达,进而促进肿瘤细胞增殖[25]。已有报道证实,在喉癌相关的辐射耐受细胞株中,通过siRNA降低PAG1的表达可以增强辐射耐受细胞的放射增敏作用[26]。LDLR和LRP1基因分别编码低密度脂蛋白受体和低密度脂蛋白受体相关蛋白1,其中LDLR基因与晚期结肠癌患者的临床分期有关[27]。而LRP1是一个多功能的具有内吞作用的细胞表面受体[28]。LRP1蛋白的表达影响着肿瘤细胞的迁移和侵袭[29],然而,对于LDLR和LRP1蛋白在放射敏感性中发挥的作用亦缺乏相关的报道。PIM1基因编码一种丝氨酸/苏氨酸激酶,研究表明,PIM1激酶参与调节细胞周期、凋亡、干细胞分化、细胞代谢和细胞自噬等细胞生理学过程,从而在肿瘤的发生发展中起重要作用[30-31]。同时,Gu等[32]证明miR-124和miR-144通过靶向PIM1可以增强前列腺癌细胞的放射敏感性。PLAUR的编码蛋白通过与Pro-UPA蛋白的相互作用,主要参与伤口愈合、血块溶解、组织重塑等生理过程;同时,进一步激活相关的促侵袭蛋白的表达,如血纤维蛋白溶酶原、间质金属蛋白酶等,导致细胞外基质降解,促进肿瘤细胞的迁移和侵袭[33]。而三阴性乳腺癌细胞的强侵袭性正是由于相关蛋白尿激酶纤维蛋白溶酶原激活物受体、泛素相关蛋白和基质金属蛋白酶的高表达造成的[34]。
本研究通过实时荧光定量PCR实验,验证潜在辐射抗性基因在5 Gy γ射线照射后的人结直肠癌HCT116细胞中的mRNA差异表达水平,结果显示,潜在抗性基因的表达趋势与测序结果一致,进一步证实了高通量测序结果的准确性。
通过对结直肠癌芯片数据的挖掘以及生物信息学分析,共筛选出NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR等6个与结肠癌细胞抗辐射能力关系密切的核心基因,这些关键基因有望成为预测结直肠癌患者的放射敏感性以及放疗疗效评价的重要基因,为接下来我们深入的研究提供参考依据和实验基础。例如,采用体内外实验模型,增加和降低这些基因的表达水平从而深入了解这些基因是否直接影响直肠癌的放射敏感性,如果是,那么这些基因的调节差异以及可能不同的作用机制何在?如果在结直肠癌是如此,在其他肿瘤的放疗敏感性的调节下是否也会产生同样的影响?总之,本研究为结直肠癌辐射抵抗的分子机制研究提供了初步的理论基础和方向,具有重要意义和临床应用价值。
应用生物信息学确定结直肠癌辐射抗性细胞的差异表达基因
Identification of genes for radiation resistance in colorectal cancer cells using bioinformatics analysis
-
摘要:
目的基于生物信息学的方法,筛选结直肠癌辐射抗性细胞中的差异表达基因,从分子水平上初步探讨与结直肠癌抗辐射相关的潜在基因。 方法从基因芯片公共数据库(GEO)中下载耐辐射的结直肠癌细胞基因表达谱数据(GSE43206),并利用R语言中的limma包进行差异基因筛选。对差异基因中的编码基因分别进行基因本体论(GO)富集分析、京都基因和基因组百科全书(KEGG)通路分析以及蛋白相互作用(PPI)分析,进一步筛选出PPI网络中的关键基因。通过实时荧光定量PCR实验确定5 Gy γ射线照射后人结肠癌HCT116细胞中关键基因的mRNA相对表达水平。采用Student t-test检验进行统计学分析,P < 0.05表示差异有统计学意义。 结果共筛选出101个差异基因,包含67个上调基因,34个下调基因。GO富集分析发现这些差异基因在细胞迁移、DNA复制等生物学过程中富集。KEGG通路分析证实这些差异基因主要富集在乏氧诱导因子1信号通路。通过构建PPI网络,筛选出NDRG1、PAG1、LRP1、PIM1、LDLR和PLAUR共6个与结直肠癌抗辐射相关的潜在基因。实时荧光定量PCR实验结果显示,与照射前比较,照射后人结肠癌HCT116细胞中NDRG1、PAG1、LRP1、PIM1、LDLR和NDRG1、PAG1、LRP1、PIM1、LDLR关键基因的mRNA表达量显著上升,差异均有统计学意义(t=49.981,P < 0.01;t=26.420、28.698、21.358、23.545,均P < 0.05;t=50.601,P < 0.01)。 结论利用生物信息学能够快速地筛选出与结直肠癌抗辐射相关的潜在基因,且潜在基因在结直肠癌HCT116细胞中差异表达。 Abstract:ObjectiveTo preliminarily explore potential genes related to radiation resistance in colorectal cancer at the molecular level, we employed bioinformatics to screen different expression genes for radiation resistance in colorectal cancer cells. MethodsThe comparison between the gene expression levels of radiation resistance colorectal cancer cell lines and parental cell lines was downloaded from the Gene Expression Omnibus(GEO) database. The differentially expressed genes(DEGs) were screened by using the R Programming Language and were analyzed through Gene Ontology(GO) functional enrichment analysis and kyoto encyclopedia of genes and genomes(KEGG) pathway analysis and by using protein-protein interaction(PPI) networks. The hub genes were obtained on the basis of a PPI network. The mRNA relative expression level of the hub genes was verified via quantitative real-time polymerase chain reaction in HCT116 after radiation. The statistical significance of the results was analyzed via student t-test. ResultsA total of 101 DEGs were found in GSE43206, including 67 upregulated genes and 34 downregulated genes. The GO enrichment analysis suggested that these DEGs are enriched in biological processes, including cell migration and DNA replication. KEGG pathway analysis indicated that these DEGs were mainly enriched in the hypoxia inducible factor-1 signaling pathway. Six radiation resistance genes with high connectivity were identified on the basis of the PPI networks, including NDRG1, PAG1, LRP1, PIM1, LDLR, and PLAUR. Quantitative real-time polymerase chain reation verified that the expression levels of hub genes were markedly up-regulated in HCT116 after radiation, including NDRG1、PAG1、LRP1、PIM1、LDLR and PLAUR (t=49.981, P < 0.01; t=26.420, 28.698, 21.358, 23.545, all P < 0.05; t=50.601, P < 0.01). ConclusionsThe use of bioinformatics enabled effectively screening radiation resistance genes in colorectal cancer, which can be used for further researches. The molecular biology experiments confirmed the differential expression of potential genes after irradiation in colorectal cancer cell HCT116. -
Key words:
- Colorectal neoplasms /
- Radiation resistance /
- Different expression genes /
- Bioinformatics
-
图 3 潜在辐射抗性基因mRNA相对表达量
Figure 3. The relative expression of potential radiation resistance genes
表 1 蛋白相互作用网络中根据节点中心度筛选的前10个基因
Table 1. The top 10 genes in protein-protein interaction network Network topology parameters
基因名 点度中心性 基因名 点度中心性 NDRG1 18 NDRG1 29.78 PAG1 16 LDLR 29 ENO2 12 HSPA5 25.95 RGS2 10 LRP1 24.87 LRP1 9 PIM1 25.03 PIM1 8 HSP90AA1 23.4 VEGFA 8 PAG1 22.57 LDLR 7 PLAUR 22.45 PLAUR 7 EFNA1 22.33 EPHA2 6 EPHA3 22.28  下载: 导出CSV
下载: 导出CSV
-
[1] Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018[J]. CA Cancer J Clin, 2018, 68(1):7-30. DOI:10.3322/caac.21442. [2] Cong XU, Cao K, Liu L, et al. Advances in sensitive biomarkers of radiotherapy in colorectal cancer patients[J]. J Transl Med, 2017, 4(9):84-87. DOI:10.3969/j.issn.2095-6894.2017.09.020. [3] 储小飞, 赵舒怡, 樊赛军.肿瘤干细胞与辐射抗性的研究进展[J].国际放射医学核医学杂志, 2015, 39(5):431-436. DOI:10.3760/cma.j.issn.1673-4114.2015.05.019.
Chu XF, Zhao SY, Fan SJ. Advance progress of cancer stem cells and radioresistance[J]. Int J Radiat Med Nucl Med, 2015, 39(5):431-436. doi: 10.3760/cma.j.issn.1673-4114.2015.05.019[4] 杨丽君, 万晓晨.结直肠癌中miR-106b增强肿瘤细胞放疗抵抗作用机制的初步分析[J].中国现代医生, 2018, 56(1):29-31, 35.
Yang LY, Wan XC. Preliminary analysis on the mechanism of miR-106b enhancing the radioresistance of tumor cells in colorectal cancer[J]. Chin Mod Doctor, 2018, 56(1):29-31, 35.[5] Clough E, Barrett T. The Gene Expression Omnibus Database[J]. Methods Mol Biol, 2016, 1418:93-110. DOI:10.1007/978-1-4939-3578-9_5. [6] He X, Fan L, Wu Z, et al. Gene expression profiles reveal key pathways and genes associated with neuropathic pain in patients with spinal cord injury[J]. Mol Med Rep, 2017, 15(4):2120-2128. DOI:10.3892/mmr.2017.6231. [7] Keerin P, Kurutach W, Boongoen T. Cluster-based KNN missing value imputation for DNA microarray data[J]. Int J Data Min Bioinform, 2012, 15(2):445-450. DOI:10.1109/ICSMC.2012. 6377764. [8] Liew AW, Law NF, Yan H. Missing value imputation for gene expression data:computational techniques to recover missing data from available information[J]. Brief Bioinform, 2011, 12(5):498-513. DOI:10.1093/bib/bbq080. [9] Ritchie ME, Phipson B, Wu D, et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies[J/OL]. Nucleic Acids Res, 2015, 43(7): e47[2018-05-02]. https: //www. ncbi. nlm. nih. gov/pubmed/25605792. DOI: 10.1093/nar/gkv007. [10] Peri S, Navarro JD, Amanchy R, et al. Development of human protein reference database as an initial platform for approaching systems biology in humans[J]. Genome Res, 2003, 13(10):2363-2371. DOI:10.1101/gr.1680803. [11] Smoot ME, Ono K, Ruscheinski J, et al. Cytoscape 2.8:new features for data integration and network visualization[J]. Bioinformatics, 2011, 27(3):431-432. DOI:10.1093/bioinformatics/btq675. [12] Shannon P, Markiel A, Ozier O, et al. Cytoscape:a software environment for integrated models of biomolecular interaction networks[J]. Genome Res, 2003, 13(11):2498-2504. DOI:10.1101/gr.1239303. [13] Lin CY, Chin CH, Wu HH, et al. Hubba:hub objects analyzer-a framework of interactome hubs identification for network biology[J]. Nucleic Acids Res, 2008, 36(Web Server issue):W438-W443. DOI:10.1093/nar/gkn257. [14] Cline MS, Smoot M, Cerami E, et al. Integration of biological networks and gene expression data using Cytoscape[J]. Nat Pro, 2007, 2(10):2366-2382. DOI:10.1038/nprot.2007.324. [15] Irizarry RA, Hobbs B, Collin F, et al. Exploration, normalization, and summaries of high density oligonucleotide array probe level data[J]. Biostatistics, 2003, 4(2):249-264. DOI:10.1093/biostatistics/4.2.249. [16] Jeong H, Mason SP, Barabási AL, et al. Lethality and centrality in protein networks[J]. Nature, 2001, 411(6833):41-42. DOI:10.1038/35075138. [17] Goh KI, Oh E, Kahng B, et al. Betweenness centrality correlation in social networks[J/OL]. Phys Rev E Stat Nonlin Soft Matter Phys, 2003, 67(1 Pt 2): 017101[2018-05-02]. https: //www. ncbi. nlm. nih. gov/pubmed/12636633. DOI: 10.1103/PhysRevE.67.017101. [18] Ronnekleiv-Kelly SM, Burkhart RA, Pawlik TM. Molecular markers of prognosis and therapeutic targets in metastatic colorectal cancer[J]. Surg Oncol, 2016, 25(3):190-199. DOI:10.1016/j.suronc.2016. 05.018. [19] Leroi N, Lallemand F, Coucke P, et al. Impacts of Ionizing Radiation on the Different Compartments of the Tumor Microenvironment[J]. Front Pharmacol, 2016, 7:78. DOI:10.3389/fphar.2016.00078. [20] Moncharmont C, Levy A, Guy JB, et al. Radiation-enhanced cell migration/invasion process:a review[J]. Crit Rev Oncol Hematol, 2014, 92(2):133-142. DOI:10.1016/j.critrevonc.2014.05.006. [21] Basu AK. DNA damage, mutagenesis and cancer[J]. Int J Mol Sci, 2018, 19(4):970-970. DOI:10.3390/ijms19040970. [22] Lee SY, Jeong EK, Ju MK, et al. Induction of metastasis, cancer stem cell phenotype, and oncogenic metabolism in cancer cells by ionizing radiation[J]. Mol Cancer, 2017, 16(1):10. DOI:10.1186/s12943-016-0577-4. [23] Sharma A, Mendonca J, Ying J, et al. The prostate metastasis suppressor gene NDRG1 differentially regulates cell motility and invasion[J]. Mol Oncol, 2017, 11(6):655-669. DOI:10.1002/1878-0261.12059. [24] Ma J, Gao Q, Zeng S, et al. Knockdown of NDRG1 promote epithelial-mesenchymal transition of colorectal cancer via NF-κB signaling[J]. J Surg Oncol, 2016, 114(4):520-527. DOI:10.1002/jso.24348. [25] Agarwal S, Ghosh R, Chen Z, et al. Transmembrane adaptor protein PAG1 is a novel tumor suppressor in neuroblastoma[J]. Oncotarget, 2016, 7(17):24018-24026. DOI:10.18632/oncotarget.8116. [26] Ke Q, Wu J, Ming B, et al. Identification of the PAG1 gene as a novel target of inherent radioresistance in human laryngeal carcinoma cells[J]. Cancer Biother Radiopharm, 2012, 27(10):678-684. DOI:10.1089/cbr.2012.1191. [27] Wang C, Li P, Xuan J, et al. Cholesterol Enhances Colorectal Cancer Progression via ROS Elevation and MAPK Signaling Pathway Activation[J]. Cell Physiol Biochem, 2017, 42(2):729-742. DOI:10.1159/000477890. [28] Lillis AP, Van Duyn LB, Murphy-Ullrich JE, et al. LDL receptor-related protein 1:unique tissue-specific functions revealed by selective gene knockout studies[J]. Physiol Rev, 2008, 88(3):887-918. DOI:10.1152/physrev.00033.2007. [29] Li Y, Reynolds RC. LRP1:a tumor and metastasis promoter or suppressor?[J]. Biochem Pharmacol, 2012, 1(5):121. DOI:10.4172/2167-0501.1000e121. [30] Xu J, Zhang T, Wang T, et al. PIM kinases:an overview in tumors and recent advances in pancreatic cancer[J]. Future Oncol, 2014, 10(5):865-876. DOI:10.2217/fon.13.229. [31] Zemskova M, Sahakian E, Bashkirova S, et al. The PIM1 kinase is a critical component of a survival pathway activated by docetaxel and promotes survival of docetaxel-treated prostate cancer cells[J]. J Biol Chem, 2008, 283(30):20635-20644. DOI:10.1074/jbc.M709479200. [32] Gu H, Liu M, Ding C, et al. Hypoxia-responsive miR-124 and miR-144 reduce hypoxia-induced autophagy and enhance radiosensitivity of prostate cancer cells via suppressing PIM1[J]. Cancer Med, 2016, 5(6):1174-1182. DOI:10.1002/cam4.664. [33] Blasi F, Carmeliet P. uPAR:a versatile signalling orchestrator[J]. Nat Rev Mol Cell Biol, 2002, 3(12):932-943. DOI:10.1038/nrm977. [34] Foekens JA, Peters HA, Look MP, et al. The urokinase system of plasminogen activation and prognosis in 2780 breast cancer patients[J]. Cancer Res, 2000, 60(3):636-643. -

 点击查看大图
点击查看大图

图(3)表(1)
计量
- 文章访问数: 3671
- HTML全文浏览量: 2396
- PDF下载量: 11