


 下载:
下载:
-
耐辐射奇球菌(Deinococcus radiodurans,DR)因其强大的辐射抗性而闻名于世,它拥有极强的电离辐射抗性,且对紫外线、干燥、氧化剂等造成的DNA损伤有强大的修复作用,因此被公认为是研究辐射抗性的模式生物[1]。就目前的研究成果来看,DR的辐射抗性机制主要包括:①特殊的生理特性与功能;②高效的DNA修复能力;③强大的抗氧化能力[2]。
-
DR是一种橘红色非致病细菌,以二联体或四联体的形式存在。从细胞壁的成分上看,DR属于革兰氏阴性菌,但是对革兰氏染色却呈阳性反应,这可能是由于它的肽聚糖层太厚,不容易脱色所致[3]。White等[4]对DR进行了全基因组测序,发现DR有冗长的基因组。DNA双链在受到外界攻击断裂后,会迅速启动同源重组修复,在此过程中需要寻找它的同源片段。DR拥有多拷贝的基因组数,这使其损伤的DNA更易找到同源的片段进行修复,从而有效抵抗损伤。且DR基因组中的GC(鸟嘌呤与胞嘧啶)含量很高,达66.6%,高的GC含量使得DNA的密度更高,也更不易变性[5]。DR的类核在生长平台期呈现环状结构,这种致密的结构即使在辐射状态下也不会发生改变,这样基因组在外界胁迫条件下可以被更好地保护起来[6]。
-
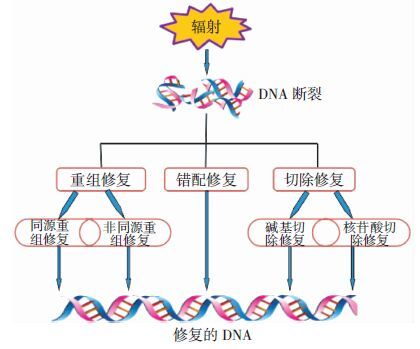
电离辐射作用于生命体,会直接造成DNA的损伤。DR与其他辐射敏感细菌一样,在极端的环境下都会产生大量的DNA碎片,但它却能在十几个小时内将产生的碎片修复而不发生突变[7]。存在于DR中的DNA修复路径主要有重组修复、错配修复和切除修复等(图 1)。同源重组修复是利用同源DNA为模板,修复损伤处的DNA。RecA是同源重组修复中的重要蛋白,它能结合双链DNA,在ATP存在的情况下形成螺旋丝,对DNA起保护作用[8]。单链退火途径是一种非依赖RecA的修复途径,通过ddrA、ddrB等蛋白的作用,在早期保护单链DNA免受降解并促进修复[9]。Zahradka等[10]发现的延伸合成依赖的单链退火途径是DR中一种更为高效的修复机制,它使具有重叠同源末端的染色体片段既可作为引物,又可作为模板,使得修复更加精准有效。错配修复主要用来纠正复制过程中产生的错配碱基。MutS能识别并结合错配的碱基位点,在ATP存在的情况下募集修复蛋白,最终在DNA聚合酶Ⅲ和DNA连接酶的作用下完成修复,从而保证了DR复制和重组的高保真性[11]。切除修复是将损伤的碱基或核苷酸进行切除从而达到DNA修复的目的。DR中包括多种与碱基切除修复相关的核酸内切酶、核酸外切酶以及糖苷水解酶,这使得这种简单高效的修复途径在DR中有效发挥作用[12]。核苷酸切除修复主要对于嘧啶二聚体的形成进行切除,DR中包括UvrABC和UvDE两条路径,研究发现,其中任何一条路径的缺失都不会影响其核苷酸切除修复功能的发挥,显示出DR修复机制的精密与高效性[13]。过多的DNA碎片堆积会对细胞产生有害作用,DR除具有高效的DNA修复能力外,还能及时地将多余的DNA碎片排出细胞,并通过细胞外核酸酶降解,防止受损的碱基重新进入细胞内参与基因组的修复,从而降低了基因的突变率[14]。

图 1 DNA修复途径简图
Figure 1. Pathway of DNA repair
-
电离辐射在直接导致生物大分子物质(DNA、RNA及蛋白质等)发生电离和激发等损失的同时,也可致使大量活性氧自由基的产生,从而对生命体产生间接损害[15]。DR能抵抗多种外界压力,其强大的抗氧化作用不容忽视。存在于DR中的抗氧化物质分为酶类和非酶类两种,酶类主要包括过氧化氢酶和超氧化物歧化酶等,其中过氧化氢酶的活性比大肠杆菌高30倍,而超氧化物歧化酶的活性比大肠杆菌高6倍[16]。非酶类物质的抗氧化作用主要与类胡萝卜素以及细胞内高的Mn2+/Fe2+值相关。DR编码一种特有的类胡萝卜素deinoxanthin[17],它是其自身呈现特殊橘红色的主要色素。所有的类胡萝卜素均含有一种特殊的共轭双键结构,这一结构决定着它的抗氧化能力[18],deinoxanthin含有13个共轭双键,使它比其他种类的类胡萝卜素具有更强的氧化抗性能力。研究发现,deinoxanthin不仅能有效清除单线态氧和过氧化物自由基,还能对DNA起保护作用,并防止蛋白质被氧化[19]。说明这一特有的色素在DR的抗性能力中发挥着重要功能。
金属离子通过结合蛋白质在生物体的生命活动中发挥重要功能。DR体内的Mn2+/Fe2+值为辐射敏感细菌的70~300倍。研究表明,Mn2+的聚集并不能防止DNA双链的断裂,它的功能在于对蛋白质的保护,使其不被氧化,并参与对活性氧自由基的清除[20]。游离的Fe2+会催化H2O2发生Fenton反应产生大量自由基,从而导致细胞的氧化损伤。在细胞受损后,自由基首要的攻击对象就是以Fe2+作为辅基的蛋白,在DR中,由于Mn2+浓度较高而Fe2+浓度较低,蛋白会倾向以Mn2+作为辅基,这样可以减少蛋白损伤。DR的整个抗氧化系统相辅相成,在抗击外界辐射、氧化等多种胁迫过程中发挥有效作用。
-
与这三大类主要抗性机制相关的还有许多DR中的特有基因,它们在其抗性能力中发挥重要作用。PprI被认为是调节多种辐射相关蛋白的开关基因,在DR受到辐射后,它可上调至少210个基因的表达,其中21个与DNA的损伤修复相关[21]。DdrO被认为是一种负性调控蛋白,它可与许多辐射抗性相关基因(如recA、ssB、pprA等)的启动子相结合,从而抑制其表达。Wang等[22]发现,在Mn2+存在的情况下,PprI蛋白能将DdrO切割,阻止它与其他基因启动子的结合,直接在转录水平上调节基因的表达。这一研究说明了PprI的重要性,也从侧面证明了高浓度的Mn2+在DR中的重要功能。PprM是一个依赖pprI的调控发挥作用的冷休克蛋白,研究表明pprM基因的缺失会导致DR的辐射抗性下降[23]。近来有报道称,pprM能调节DR中过氧化氢酶KatE1的表达,凸显了pprM在其氧化抗性中的地位[24]。李伟[25]对PprM蛋白的结构进行分析,发现它拥有DNA和RNA 结合位点,并能与编码组氨酸激酶DR_A0355的mRNA 区域相互作用,猜测pprM可能是潜在的结合RNA的辐射应激开关。
-
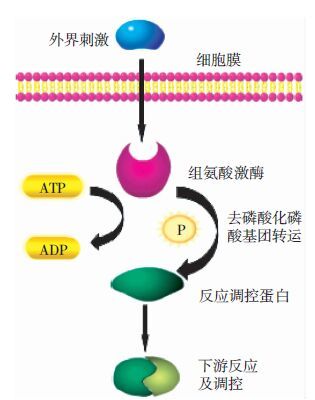
组氨酸激酶是细菌信号转导系统TCS的重要组成部分,它在细菌抗击外界胁迫作用的过程中发挥着重要作用[26]。1986年研究大肠杆菌时首次发现TCS[27]。经典的TCS包括一个组氨酸激酶和一个与之协作的响应调节因子。在接受外界刺激后,组氨酸激酶在组氨酸残基上自动磷酸化,然后将这个磷酰基转移到它相应的反应调节因子上,从而调控其下游的表达和功能。组氨酸激酶包括两个结构域,一个N末端的磷酸转导结构域DHp和一个C末端的催化以及结合ATP 的结构域CA。而反应调控因子控制着信号的输出功能,它有一个N端的信号接收结构域和一个C端的信号输出结构域,通常有一个相对保守区域来催化磷酸基团的转运[28](TCS通路简图见图 2)。随着TCS研究的深入,多组份系统也随之被发现。许多生物中存在一个组氨酸激酶和多个反应调节器以及多个组氨酸激酶和一个反应调节器组成TCS的情况。在结核分支杆菌中,组氨酸激酶1和组氨酸激酶2相互作用,然后作用于同一反应调节因子TcrA,从而触发下级反应[29]。在海藻中则存在组氨酸激酶Hik2与两个反应调控蛋白Rre1和RppA组成TCS的情况[30],这说明了TCS形式的多样,也为细菌TCS的进一步研究提供了新思路。
DR拥有如此强大的生命力,必然是有复杂的网络调控系统使其在各种逆性环境中迅速作出反应与调节的,而TCS这一信号转导系统也同样存在于DR中。Makarova等[31]在2001年报道了DR有23个编码组氨酸激酶的基因和29个编码响应调节因子的基因。郝艳华[32]研究了DtrA/DtrB这一TCS在DR中的作用,研究结果表明DtrA/DtrB的双缺失导致DR的热效应灵敏度升高,但却能增强其抗紫外的能力。RadS/RadR组成的TCS与DR的辐射抗性息息相关,其中RadS是一种组氨酸激酶,RadR是一种响应调节因子,两者位于同一个操纵序列,研究结果表明RadS/RadR的单突变株和双突变株都表现出辐射敏感性增加[33]。这些研究都说明了TCS在DR应对极端环境中发挥重要作用,对进一步了解DR的抗性机制及信号通路也有重要意义。

图 2 双组份系统信号通路简图
Figure 2. Two component system signal transduction pathway
-
我们致力于DR辐射抗性的研究,目的在于将它强大的抗辐射能力应用于实践。在福岛核事故发生后,人们对于核污染的处理更为重视。DR能作为生物吸附剂吸附溶于水的铀[34],同时它含有的NiCoT基因能净化核反应堆中的微量钴,这对于核废料的处理无疑是一大福音[35]。Dalmaso等[36]将DR培养在含有人的毛发和鸡毛的培养基中,发现其不仅能生长,而且能促进角蛋白酶的表达,从而降解角蛋白,这能让它在环境可持续发展中做出贡献。在农业方面,DR已应用于增加酵母酿酒产量、提高乳杆菌的乳酸产量等[37-38]。在临床医药研究方面,DR对于肿瘤的治疗以及核辐射的防护作用的研究也有着越来越多突破。研究人员将PprI蛋白提取后直接注入受辐照的小鼠体内,经比对发现γ射线对小鼠的急性损伤程度显著下降[39]。类胡萝卜素自身能清除氧自由基,并能与维生素C和维生素E等其他的抗氧化物质相互协同[40],使其抗氧化能力得到更大地发挥,它能增强人体免疫力,抑制肿瘤的发展。研究人员将DR中的类胡萝卜素deinoxanthin转入细胞后,发现其能诱导肿瘤细胞的凋亡[41]。而将DR中富含Mn2+、无机磷酸盐、肽类、氨基酸的细胞提取物应用于人外周血白血病T细胞后,发现它能提高细胞受辐照后的存活率[42]。这些对于DR的应用有着重要意义,也为我们了解DR提供了多方面的思路。DR的强大抗性能力是各个网络调控系统相互作用、相互协调的结果,各方面机制的研究仍有待进一步深入,相信我们更深入的研究会让其应用的领域更为广泛。
耐辐射奇球菌辐射抗性机制及其双组份系统的研究
The mechanism of resistance and two-component system in Deinococcus radiodurans
-
摘要: 耐辐射奇球菌拥有极强的辐射抗性能力并能在多种极端环境下存活。其特有的一些辐射抗性基因在其抗性机制中发挥重要作用。双组份系统是广泛存在于细菌中的一种信号转导系统,它能协助细菌应对外界环境的变化,对维持各种生命活动至关重要。双组份系统作为一种应激响应机制在耐辐射奇球菌中有重要作用。笔者就耐辐射奇球菌的抗性机制及其双组份系统的研究进行综述。Abstract: Deinococcus radiodurans(DR) has strong resistance to radiation and comprises a series of mechanisms that allow it to survive under various extreme conditions. Some of the genes existing in DR particularly contribute to its robust viability. Two-component system(TCS) is a signal transduction system that widely exists in bacteria. This system can assist the bacteria in coping with the change in external environment and is vital for various life activities. Given that TCS plays an important role in resistance mechanism as a stress response system, this article reviewed the resistant mechanism and the TCS in DR.
-
Key words:
- Deinococcus /
- Radiation tolerance /
- Two-component system
-
[1] Cox MM, Battista JR.Deinococcus radiodurans-the consummate survivor[J].Nat Rev Microbiol, 2005, 3(11):882-892.DOI:10.1038/nrmicro1264. [2] Munteanu A, Uivarosi V, Andries A.Recent progress in understanding the molecular mechanisms of radioresistance in Deinococcus bacteria[J].Extremophiles, 2015, 19(4):707-719.DOI:10.1007/s00792-015-0759-9. [3] Work E, Griffiths H.Morphology and chemistry of cell walls of Micrococcus radiodurans[J].J Bacteriol, 1968, 95(2):641-657. [4] White O, Eisen JA, Heidelberg JF, et al.Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1[J].Science, 1999, 286(5444):1571-1577. doi: 10.1126/science.286.5444.1571 [5] Long H, Kucukyildirim S, Sung W, et al.Background mutational features of the Radiation-Resistant bacterium deinococcus radiodurans[J].Mol Biol Evol, 2015, 32(9):2383-2392.DOI:10.1093/molbev/msv119. [6] Levin-Zaidman S, Englander J, Shimoni E, et al.Ringlike structure of the Deinococcus radiodurans genome:a key to radioresistance?[J].Science, 2003, 299(5604):254-256.DOI:10.1126/science.1077865. [7] Daly MJ.A new perspective on radiation resistance based on Deinococcus radiodurans[J].Nat Rev Microbiol, 2009, 7(3):237-245.DOI:10.1038/nrmicro2073. [8] Pobegalov G, Cherevatenko G, Alekseev A, et al.Deinococcus radiodurans RecA nucleoprotein filaments characterized at the single-molecule level with optical tweezers[J].Biochem Biophys Res Commun, 2015, 466(3):426-430.DOI:10.1016/j.bbrc.2015.09.042. [9] Sugiman-Marangos SN, Weiss YM, Junop MS.Mechanism for accurate, protein-assisted DNA annealing by Deinococcus radiodurans DdrB[J].Proc Natl Acad Sci U S A, 2016, 113(16):4308-4313.DOI:10.1073/pnas.1520847113. [10] Zahradka K, Slade D, Bailone A, et al.Reassembly of shattered chromosomes in Deinococcus radiodurans[J].Nature, 2006, 443(7111):569-573.DOI:10.1038/nature05160. [11] Zhang H, Xu Q, Lu M, et al.Structural and functional studies of MutS2 from Deinococcus radiodurans[J/OL].DNA Repair(Amst), 2014, 21:111-119[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/24811920.DOI:10.1016/j.dnarep.2014.04.012. [12] Agapov AA, Kulbachinskiy AV.Mechanisms of stress resistance and gene regulation in the radioresistant bacterium deinococcus radiodurans[J].Biochemistry(Mosc), 2015, 80(10):1201-1216.DOI:10.1134/S0006297915100016. [13] Tanaka M, Narumi I, Funayama T, et al.Characterization of pathways dependent on the uvsE, uvrA1, or uvrA2 gene product for UV resistance in Deinococcus radiodurans[J].J Bacteriol, 2005, 187(11):3693-3697.DOI:10.1128/JB.187.11.3693-3697.2005. [14] Wang L, Tan H, Cheng K, et al.Sec pathway influences the growth of Deinococcus radiodurans[J].Curr Microbiol, 2015, 70(5):651-656.DOI:10.1007/s00284-014-0767-5. [15] Slade D, Radman M.Oxidative stress resistance in Deinococcus radiodurans[J].Microbiol Mol Biol Rev, 2011, 75(1):133-191.DOI:10.1128/MMBR.00015-10. [16] Wang P, Schellhorn HE.Induction of resistance to Hydrogen peroxide and radiation in Deinococcus radiodurans[J].Can J Microbiol, 1995, 41(2):170-176. doi: 10.1139/m95-023 [17] Tian B, Xu Z, Sun Z, et al.Evaluation of the antioxidant effects of carotenoids from Deinococcus radiodurans through targeted mutagenesis, chemiluminescence, and DNA damage analyses[J].Biochim Biophys Acta, 2007, 1770(6):902-911.DOI:10.1016/j.bbagen.2007.01.016. [18] Woodall AA, Britton G, Jackson MJ.Carotenoids and protection of phospholipids in solution or in liposomes against oxidation by peroxyl radicals:relationship between carotenoid structure and protective ability[J].Biochim Biophys Acta, 1997, 1336(3):575-586. doi: 10.1016/S0304-4165(97)00007-X [19] Tian B, Sun Z, Shen S, et al.Effects of carotenoids from Deinococcus radiodurans on protein oxidation[J].Lett Appl Microbiol, 2009, 49(6):689-694.DOI:10.1111/j.1472-765X.2009.02727.x. [20] Daly MJ, Gaidamakova EK, Matrosova VY, et al.Accumulation of Mn(II) in deinococcus radiodurans facilitates gamma-radiation resistance[J].Science, 2004, 306(5698):1025-1028.DOI:10.1126/science.1103185. [21] Lu H, Gao G, Xu G, et al.Deinococcus radiodurans PprI switches on DNA damage response and cellular survival networks after radiation damage[J].Mol Cell Proteomics, 2009, 8(3):481-494.DOI:10.1074/mcp.M800123-MCP200. [22] Wang Y, Xu Q, Lu H, et al.Protease activity of PprI facilitates DNA damage response:Mn2+-dependence and substrate sequence-specificity of the proteolytic reaction[J/OL].PLoS One, 2015, 10(3):e0122071[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/25811789.DOI:10.1371/journal.pone.0122071. [23] Ohba H, Satoh K, Sghaier H, et al.Identification of PprM:a modulator of the PprI-dependent DNA damage response in Deinococcus radiodurans[J].Extremophiles, 2009, 13(3):471-479.DOI:10.1007/s00792-009-0232-8. [24] Jeong SW, Seo HS, Kim MK, et al.PprM is necessary for up-regulation of katE1, encoding the major catalase of Deinococcus radiodurans, under unstressed culture conditions[J].J Microbiol, 2016, 54(6):426-431.DOI:10.1007/s12275-016-6175-8. [25] 李伟.耐辐射奇球菌PprM蛋白相互作用RNA的筛选及鉴定[D].衡阳:南华大学, 2015.
Li W.The research on screening and identifying of PprM target RNAs in deinococcus radiodurans[D].Hengyang:University of south China, 2015.[26] Zschiedrich CP, Keidel V, Szurmant H.Molecular mechanisms of Two-Component signal transduction[J].J Mol Biol, 2016, 428(19):3752-3775.DOI:10.1016/j.jmb.2016.08.003. [27] Nixon BT, Ronson CW, Ausubel FM.Two-component regulatory systems responsive to environmental stimuli share strongly conserved domains with the Nitrogen assimilation regulatory genes ntrB and ntrC[J].Proc Natl Acad Sci U S A, 1986, 83(20):7850-7854. doi: 10.1073/pnas.83.20.7850 [28] Bourret RB.Receiver domain structure and function in response regulator proteins[J].Curr Opin Microbiol, 2010, 13(2):142-149.DOI:10.1016/j.mib.2010.01.015. [29] Shrivastava R, Ghosh AK, Das AK.Probing the nucleotide binding and phosphorylation by the histidine kinase of a novel three-protein two-component system from Mycobacterium tuberculosis[J].FEBS Lett, 2007, 581(9):1903-1909.DOI:10.1016/j.febslet.2007.03.089. [30] Ibrahim IM, Puthiyaveetil S, Allen JF.A Two-Component regulatory system in transcriptional control of photosystem stoichiometry:Redox-Dependent and Sodium Ion-Dependent phosphoryl transfer from cyanobacterial histidine kinase Hik2 to response regulators Rre1 and RppA[J/OL].Front Plant Sci, 2016, 7:137[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/26904089.DOI:10.3389/fpls.2016.00137. [31] Makarova KS, Aravind L, Wolf YI, et al.Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics[J].Microbiol Mol Biol Rev, 2001, 65(1):44-79.DOI:10.1128/MMBR.65.1.44-79.2001. [32] 郝艳华.耐辐射异常球菌类双组分蛋白DtrA/DtrB在高温及UV胁迫中的作用[D].中国农业科学院, 2012.
Hao YH.The role of DtrA/DtrB in the high temperature and UV stress of the radiation resistant abnormal Streptococcus[D].Chinese Academy of Agricultural Sciences, 2012.[33] Desai SS, Rajpurohit YS, Misra HS, et al.Characterization of the role of the RadS/RadR two-component system in the radiation resistance of Deinococcus radiodurans[J].Microbiology, 2011, 157(Pt10):2974-2982.DOI:10.1099/mic.0.049361-0. [34] Yang J, Dong FQ, Dai QW, et al.Biosorption of Radionuclide Uranium by Deinococcus radiodurans[J].Guang pu xue yu guang pu fen xi, 2015, 35(4):1010-1014. [35] Gogada R, Singh SS, Lunavat SK, et al.Engineered deinococcus radiodurans R1 with NiCoT genes for bioremoval of trace Cobalt from spent decontamination solutions of nuclear power reactors[J].Appl Microbiol Biotechnol, 2015, 99(21):9203-9213.DOI:10.1007/s00253-015-6761-4. [36] Dalmaso GZ, Lage CA, Mazotto AM, et al.Extracellular peptidases from Deinococcus radiodurans[J].Extremophiles, 2015, 19(5):989-999.DOI:10.1007/s00792-015-0773-y. [37] Ma R, Zhang Y, Hong H, et al.Improved osmotic tolerance and ethanol production of ethanologenic Escherichia coli by IrrE, a global regulator of radiation-resistance of Deinococcus radiodurans[J].Curr Microbiol, 2011, 62(2):659-664.DOI:10.1007/s00284-010-9759-2. [38] Dong X, Tian B, Dai S, et al.Expression of PprI from Deinococcus radiodurans Improves Lactic Acid Production and Stress Tolerance in Lactococcus lactis[J/OL].PLoS One, 2015, 10(11):e0142918[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/20959988.DOI:10.1371/journal.pone.0142918. [39] 陈洁, 张永芹, 毕洁静, 等.耐辐射球菌PprI蛋白对急性放射损伤小鼠防护作用的初步研究[J].辐射研究与辐射工艺学报, 2012, 30(5):286-290.
Chen J, Zhang YQ, Bi JJ, et al.Studying the protective effect of deinococcus radiodurans PprI on acute radiation injury in mice[J].J Radiat Res Radiat Process, 2012, 30(5):286-290.[40] Jomova K, Valko M.Health protective effects of carotenoids and their interactions with other biological antioxidants[J/OL].Eur J Med Chem, 2013, 70:102-110[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/24141200.DOI:10.1016/j.ejmech.2013.09.054. [41] Choi YJ, Hur JM, Lim S, et al.Induction of apoptosis by deinoxanthin in human cancer cells[J].Anticancer Res, 2014, 34(4):1829-1835. [42] Daly MJ, Gaidamakova EK, Matrosova VY, et al.Small-molecule antioxidant proteome-shields in Deinococcus radiodurans[J/OL].PLoS One, 2010, 5(9):e12570[2016-10-01].http://www.ncbi.nlm.nih.gov/pubmed/20838443.DOI:10.1371/journal.pone.0012570. -

 点击查看大图
点击查看大图
图(2)
计量
- 文章访问数: 3953
- HTML全文浏览量: 3153
- PDF下载量: 4