


 下载:
下载:


-
B细胞易位基因2(B-cell translocation gene 2,BTG2)是细胞增殖抑制蛋白家族TOB/BTG的成员之一。目前的研究发现,BTG2参与包括肿瘤细胞在内的细胞分化、发育、凋亡等功能调节[1-6]。最近研究发现,人为提高BTG2的表达可以提高人乳腺癌T-47D细胞的放射敏感性,且可能与诱导细胞凋亡有关[7]。本研究通过提高和降低肿瘤细胞中BTG2的表达水平进一步了解BTG2在肿瘤细胞放射敏感性调节中的关键作用及其机制,为BTG2临床肿瘤放射治疗增敏性的预测和预后可行性提供实验基础和理论依据。
-
人乳腺癌细胞系MCF-7、MDA-MB-231,人肺癌细胞系H2170、A549、NIH-H460、SK-MES-1、HTB-58、H23和H441以及正常人上皮细胞系184A1购自美国细胞收藏中心,由苏州大学医学部放射医学与公共卫生学院肿瘤放射生物学实验室培养和保存。乳腺癌和肺癌细胞于含10%胎牛血清的DMEM(Dulbecco's modified eagle medium)培养液(美国Gibco公司)中常规培养,184A1细胞于含10%胎牛血清以及10 ng/ml人上皮细胞生长因子的MEGM(mammary epithelium basal medium)培养液(美国Clonetics公司)中培养。所有细胞都培养于37 ℃、5% CO2培养箱内。
-
全长BTG2 cDNA或全长BRCA1 cDNA定向分别装载至真核细胞表达载体pcDNA3(美国Invitrogen公司)中,重组质粒由美国Invitrogen公司进行核酸序列测定。采用脂质体介导的转染方法,将构建好的pcDNA3-BTG2质粒导入部分乳腺癌细胞中;同时将pcDNA3“空”质粒导入另外一部分乳腺癌细胞中作为阴性对照。将转染后的细胞在含有nemycin(G418,500 g/ml)(美国Gibco公司)的常规培养液中进一步培养,连续培养15 d后,收集所有存活克隆并汇集在一起成为稳定细胞株,继续在含nemycin(G418,500 g/ml)的培养液中培养传代。照射前将转染后的细胞在无nemycin的培养液中培养24 h。为研究pcDNA3-BRCA1对BTG2调节肿瘤细胞放射敏感性的影响,同时在含nemycin的培养液中将pcDNA3-BRCA1质粒瞬时转染到已转染了pcDNA3-BTG2质粒的MCF-7或MDA-MB-231稳定细胞株中,转染后24 h进行照射并完成细胞生存实验。
-
收集指数期生长的细胞,采用137Cs γ射线照射(照射源由美国Georgetown大学辐照中心提供),吸收剂量率为0.8 Gy/min,源靶距为73.5 cm,照射视野为30 cm×30 cm。
-
将1.0×105对数期生长的细胞接种于24孔培养板,采用lipofectamine 2000(美国Invitrogen公司)转染试剂将siRNA-BRCA1和“空”siRNA转染至细胞中,转染基本操作则按照说明书进行。转染后于48 h时采用胰酶消化收取细胞,然后照射完成噻唑蓝实验。
-
将细胞以每孔3×105个/ml的细胞接种于96孔细胞培养板中,在30 ℃、5%CO2细胞培养箱中培养48 h后加入10 μl噻唑蓝(10 mg/ml),继续培养4 h,800 g离心5 min(1000 r/min,离心半径为105 mm),弃上清,每孔加入100 μl二甲基亚砜(美国Sigma公司),振动10 min,酶标仪测定570 nm处的光密度。根据光密度算出细胞增殖抑制率。增殖抑制率=[1-(实验组光密度-调零孔光密度)/(对照孔光密度-调零孔光密度)]×100%。
-
将需要照射的各组细胞按不同照射剂量接种不同细胞数接种到60 mm培养皿中,培养3周,用甲醇固定,加姬姆萨应用液(美国Sigma公司)染色30 min,流水洗净,计数细胞数≥50的克隆个数。克隆形成率=(对照组克隆数/实验组细胞数)×100%,存活分数=克隆数/[实验组细胞数×(观察的克隆数/接种的细胞数)]。
-
将收集的细胞样品加裂解液于冰上裂解细胞,离心10 min(10 000 r/min,离心半径为105 mm),取上清液,二喹啉甲酸(bicinchoninic acid,BCA)法蛋白定量试剂盒(美国Invitrogen公司)调整蛋白至相同浓度。样品加入等体积2×十二烷基硫酸钠凝胶加样缓冲液,沸水中煮沸5 min,经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳后转膜;封闭液封闭2 h;加用封闭液稀释的BTG2(sc-30342,1:250稀释)、抗BRCA1(sc-642,1:500稀释)和抗α-Actin(sc-32252,1:2000稀释)抗体(美国Santa Cruz生物技术公司),4℃孵育过夜;用tris-buffered saline and tween 20缓冲液(简称TBST缓冲液)冲洗3次,每次15 min;加封闭液稀释相应的山羊抗鼠(sc-2060,1:2000稀释)或山羊抗兔(sc-2004,1:2000稀释)的二抗(美国Santa Cruz生物技术公司),室温孵育1 h,TBST缓冲液冲洗3次,每次15 min;用增强化学发光法检测蛋白表达状况。
-
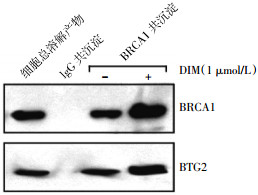
指数期生长的184A1细胞分别在含有和不含有1 μmol/L的3,3二吲哚甲烷(3,3-diindolylmethane,DIM)培养液中培养24 h,然后用冰冷的生理盐水洗涤贴壁生长的184A1细胞两次,加入0.5 ml细胞裂解液,并收集细胞,离心5 min(1000 r/min,离心半径105 mm),转移上清至新离心管中,加蛋白A珠子(美国Sigma公司)并在4 ℃下放置10 min,测定蛋白浓度。加入250 μl的免疫球蛋白G抗体(美国Santa Cruz生物技术公司)或BRCA1抗体(美国Santa Cruz生物技术公司)至500 μl的细胞裂解液中,并孵肓4 h。离心1 min(4000 r/min,离心半径105 mm)收集珠子,弃掉上清,并用细胞缓冲液冲洗3次。将珠子重新悬于60 ml 2 ×蛋白上样缓冲液中,离心1 min(4000 r/min,离心半径105 mm),弃掉珠子。行Western blot检测上清中BRCA1和BTG2蛋白的表达水平。
-
采用SPSS13.0统计学软件对数据进行统计学分析,计量资料采用均数±标准差($\bar x \pm s$)表示,组间比较采用t检验,P < 0.05表示差异有统计学意义。
-
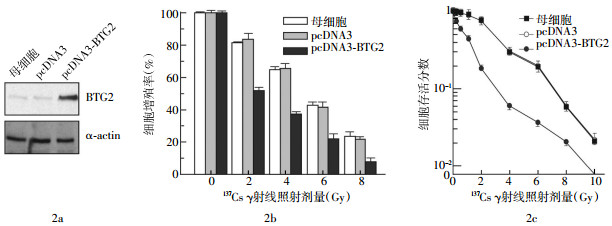
Western blot实验结果显示,在BTG2蛋白表达水平相对比较低的人乳腺癌MCF-7和MDA-MB-231细胞中,转染了“空”pcDNA3质粒的MCF-7和MDA-MB-231细胞的BTG2蛋白水平并未见任何改变;转染了pcDNA3-BTG2质粒的MCF-7和MDA-MB-231细胞的BTG2蛋白表达水平明显提高;无论是在转染了pcDNA3的质粒细胞中还是在转染了pcDNA3-BTG2的质粒细胞中,作为内参的α-actin蛋白表达水平均无明显改变,表明各组细胞总蛋白上样量基本一致(图 1a、图 2a)。这些实验结果表明,在pcDNA3-BTG2转染的细胞中BTG2蛋白水平的高表达是由于转染了BTG2所致。

图 1 B细胞易位基因2对人乳腺癌MCF-7细胞放射敏感性的影响

图 2 B细胞易位基因2对人乳腺癌MDA-MB-231细胞放射敏感性的影响
噻唑蓝实验和细胞克隆形成实验结果表明,受到不同剂量的γ射线照射后,与MCF-7、MDA-MB-231母细胞和分别转染了“空”pcDNA3质粒的MCF-7、MDA-MB-231细胞株相比,转染了pcDNA3-BTG2质粒的MCF-7和MDA-MB-231细胞株的存活率明显下降(图 1b、1c、2b、2c)。在MCF-7细胞中,IC50(导致50%细胞死亡的辐射剂量)在pcDNA3转染的细胞和pcDNA3-BTG2转染的细胞中分别为1.2 Gy以及0.3 Gy,它们之间的差异有统计学意义(t=3.12,P < 0.05)。在MDA-MB-231细胞中,IC50在pcDNA3转染的细胞和pcDNA3-BTG2转染的细胞中分别为2.8 Gy以及0.8 Gy,它们之间的差异同样有统计学意义(t=3.39,P < 0.05)。
采用噻唑蓝比色法检测不同细胞株的生长状况,结果发现,与MCF-7和MDA-MB-231母细胞相比,BTG2高表达对MCF-7和MDA-MB-231肿瘤细胞的生长并不产生明显的抑制作用。例如,pcDNA3转染的MCF-7细胞倍增时间为20 h,pcDNA3-BTG2转染的MCF-7细胞倍增时间为19 h;pcDNA3转染的MDA-MB-231细胞倍增时间为25 h,pcDNA3-BTG2转染的MDA-MB-231细胞倍增时间为24 h。此结果说明BTG2高表达对肿瘤细胞的放射敏感性的影响并不是由于BTG2表达改变影响细胞生长所致。
-
为了解BTG2调节肿瘤细胞放射敏感性的作用机制,我们前期的研究采用基因芯片法比较了pcDNA3转染的MCF-7细胞和pcDNA3-BTG2转染的MCF-7细胞内在基因表达水平,结果发现一种DNA损伤修复蛋白BRCA1的表达水平有明显改变。此次的IP-Western blot实验结果发现,BTG2与BRCA1蛋白之间形成了一个复合体。BTG2蛋白存在于采用BRCA1抗体的IP蛋白复合体中,而且DIM——BRCA1表达诱导剂明显增加了BTG2蛋白与BRCA1蛋白的结合量(图 3)。显然,这些实验结果表明BTG2蛋白与BRCA1蛋白之间可以相互结合。

图 3 采用3,3二吲哚甲烷处理184A1细胞24 h后免疫共沉淀-Western blot的检测结果
-
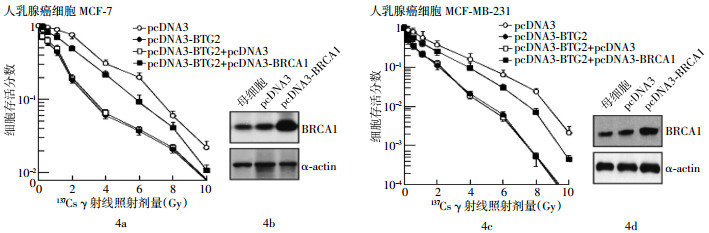
在转染了pcDNA3-BTG2质粒的细胞中同时转染pcDNA3-BRCA1或siRNA-BRCA1,采用细胞克隆形成法测定其放射敏感性,结果发现,转染了pcDNA3-BRCA1后,高表达BTG2所导致的细胞放射敏感性提高明显受到抑制(图 4)。只转染了pcDNA3-BTG2的MCF-7细胞的IC50为0.3 Gy,而共转染了pcDNA3-BTG2和pcDNA3-BRCA1的MCF-7细胞的IC50则为1.0 Gy,两者之间的差异有统计学意义(t=3.98,P < 0.05)。只转染了pcDNA3-BTG2的MDA-MB-231细胞的IC50为0.8 Gy,而共转染了pcDNA3-BTG2和pcDNA3-BRCA1的MDA-MB-231细胞的IC50则为1.9 Gy,两者之间的差异也有统计学意义(t=2.93,P < 0.05)。但是,共转染siRNA-BRCA1则导致了pcDNA3-BTG2转染细胞的放射敏感性进一步提高(图 5)。只转染了pcDNA3-BTG2的MCF-7细胞的IC50为0.3 Gy,而共转染了pcDNA3-BTG2和sirRNA-BRCA1的MCF-7细胞的IC50则为0.25 Gy(t=2.26,P < 0.05)。只转染了pcDNA3-BTG2的MDA-MB-231细胞的IC50为0.8 Gy,而共转染了pcDNA3-BTG2和siRNA-BRCA1的MDA-MB-231细胞的IC50则为0.6 Gy(t=1.97,P < 0.05)。这些实验结果表明,BRCA1在BTG2调节肿瘤放射敏感性中可能起到了非常重要的作用。

图 4 乳腺癌易感基因1高表达对B细胞易位基因2调节乳腺癌细胞放射敏感性的影响

图 5 乳腺癌易感基因1低表达对B细胞易位基因2调节乳腺癌细胞放射敏感性的影响
-
我们比较了未作任何处理的指数期生长的7种肺癌细胞系中的BRCA1和BTG2的表达水平,结果发现,BRCA1蛋白表达水平分别为:H2170细胞最高,其次为NIH-H460和SK-MES-1细胞,A549、HTB-58、H23和H441细胞中BRCA1蛋白表达水平则非常低;而BTG2的表达正好相反,在A549、HTB-58、H23和H441细胞中的表达水平比较高,而在NIH-H460和SK-MES-1细胞中的表达水平较低,在H2170细胞中则几乎无法检测到(图 6)。当采用5 Gy单剂量γ射线照射这些肺癌细胞系后24 h完成噻唑蓝实验,结果发现这7种肺癌细胞系的放射敏感性顺序为A549>HTB-58>H23>H441>SK-MES-1>NCI-H460>H2170。结合蛋白水平的表达结果可以看出,这些被检测的肺癌细胞的放射敏感性与其BRCA1的表达水平成反比,与BTG2的表达水平成正比。因此,当细胞含有高BTG2表达,低BRCA1表达时,其放射敏感性相对比较高。

图 6 5 Gy单次137Cs γ射线照射7种肺癌细胞24 h后的细胞增殖率(6a)和指数生长期的7种肺癌细胞中BTG2和BRCA1蛋白表达水平的Western blot检测结果(6b)
-
放射治疗是包括乳腺癌和肺癌在内的不同肿瘤的重要治疗手段之一,了解影响肿瘤的放射敏感性基因的改变,深入并有的放矢地研究从而提高放疗疗效一直是放射治疗基础和临床研究领域的重要课题。
BTG2蛋白是细胞增殖抑制家族TOB/BTG的成员之一,其位于人类染色体的1q32区,mRNA的长度为2717碱基对,编码一个由158个氨基酸组成的蛋白质[1]。国内外已有研究报道,BTG2蛋白作为瞬时早期反应蛋白,在包括肿瘤细胞在内的细胞分化、抑制细胞增殖和诱导细胞凋亡等方面具有重要功能[2-6]。我们以前的研究发现,BTG2高表达可以影响乳腺癌T-47细胞的放射敏感性[7]。本研究延伸了这项研究,发现BTG2高表达也可以提高乳腺癌MCF-7和MDA-MB-231细胞的放射敏感性,而且相对于MCF-7细胞(p53基因野生型的细胞)来说,BTG2对放射治疗不敏感的MDA-MB-231细胞(p53基因畸变型的细胞)的放射敏感性的提高程度更大。这在临床上具有更重要的意义,因为70%以上的中晚期乳腺癌患者的肿瘤细胞都发生了p53基因畸变,变得对放射治疗不敏感。因此,肿瘤细胞BTG2的检测可能对放射治疗的疗效和预后具有一定的意义和临床价值。目前,我们正在进一步研究BTG2调节乳腺癌放射治疗的敏感性是否是p53依赖性。
通过前期的基因芯片实验研究发现,BTG2高表达对细胞中BRCA1的表达水平有明显的影响。采用IP-Western blot方法发现BTG2蛋白与BRCA1蛋白之间确实存在相互作用,并形成了一个复合体。而且增加BRCA1的表达水平明显抑制了BTG2对肿瘤细胞放射敏感性的影响,降低BRCA1的表达水平明显增加了BTG2对肿瘤细胞放射敏感性的影响。更进一步的研究发现,BTG2与肺癌放射敏感性成正相关,而BRCA1表达水平与肺癌放射敏感性成负相关。肿瘤抑制基因BRCA1是DNA损伤修复的重要蛋白,同时也是一种抗氧化基因,提高细胞BRCA1表达水平可以明显地防护细胞对抗过氧化氢所导致的损伤[8]。这些结果为深入研究BTG2在提高肿瘤放射敏感性机制中的作用提供了重要的方向,BTG2是否也直接参与DNA损伤修复或通过BRCA1间接地参与辐射损伤修复有待进一步的研究和探讨。
总之,本研究实验结果表明,提高BTG2表达水平可以明显地提高人乳腺癌细胞的放射治疗敏感性,其作用机制可能与BRCA1蛋白的直接结合有一定的关系。这些研究也为我们深入了解BTG2作为一种新放射增敏蛋白在肿瘤放射治疗中的功能提供了研究基础和理论依据。
B细胞易位基因2的表达水平对肿瘤细胞放射敏感性的影响
Effect of B-cell translocation gene 2 alteration on radiosensitivity of cancer cells
-
摘要:
目的 研究B细胞易位基因2(BTG2)表达水平的改变对肿瘤细胞放射敏感性的影响。 方法 通过pcDNA3-BTG2脂质体转染的方法提高细胞的BTG2的表达水平,利用噻唑蓝和细胞克隆形成实验研究细胞放射敏感性的改变,应用Western blot方法研究蛋白表达水平的变化。 结果 噻唑蓝和细胞克隆形成实验结果显示,在不同剂量的γ射线照射后,提高BTG2的表达水平可明显提高乳腺癌MCF-7和MDA-MB-231细胞的放射敏感性。免疫共沉淀-Western blot实验结果显示BTG2蛋白与DNA损伤修复和抗氧化蛋白乳腺癌易感基因1(BRCA1)相互作用,形成复合物。高表达的BRCA1明显地抑制了BTG2高表达对乳腺癌细胞放射敏感性的调节作用,而降低BRCA1的表达水平则提高了BTG2对乳腺癌细胞放射敏感性的调节作用。另外,肺癌细胞放射敏感性与其所含的BRCA1的表达水平成反比,而与BTG2的表达水平成正比。 结论 BTG2的高表达明显提高了肿瘤细胞的放射敏感性,其机制可能与其同BRCA1形成复合物有关。 Abstract:Objective To investigate the effects of B-cell translocation gene 2(BTG2) overexpression on the radiosensitivity of cancer cells. Methods Cancer cells with overexpression of BTG2 were established via stable transfection of full-length human BTG2 cDNA which was inserted into a mammalian expression plasmid pcDNA3(pcDNA3-BTG2). Cell survival was determined by thiazolyl blue tetrazolium bromide (MTT) and clonogenic survival assays. Protein-protein interaction was performed by immune precipitation(IP)-Western blot assay. Protein expression was assayed by Western blot assay. Results As demonstrated in MTT assay and clonogenic survival assay, enforced expression of BTG2 significantly enhanced radiosenstivity of human breast cancer MCF-7 and MAD-MB-231 cells. The BTG2 protein was able to be determined in the breast cancer susceptibility gene1(BRCA1) IP. Silence of BRCA1 enhanced the increased radiosensitivity by BTG2, however, co-overexpression of BRCA1 reduced the BTG2-mediated radiosenstivity. Finally, the radiosensitivity of lung cancer cell lines tested exhibited a positive relationship with the levels of BTG2 protein expression and a negative correlation with the levels of BRCA1 protein expression. Conclusion The present study further demonstrates that there is a significant relationship of radiosenstivity with BTG2 and BRCA1 expression, suggesting that BTG2 may be a new and important target in cancer radiotherapy via its binding to BRCA1. -
Key words:
- Neoplasms /
- Radiation tolerance /
- B-cell translocation gene 2 /
- Gene, BRCA1
-
[1] Winkler GS. The mammalian anti-proliferative BTG/Tob protein family. J Cell Physiol, 2010, 222(1):66-72. doi: 10.1002/jcp.21919 [2] Möllerström E, Kovács A, Lövgren K, et al. Up-regulation of cell cycle arrest protein BTG2 correlates with increased overall survival in breast cancer, as detected by immunohistochemistry using tissue microarray. BMC Cancer, 2010, 10:296. doi: 10.1186/1471-2407-10-296 [3] Cullmann C, Hoppe-Seyler K, Dymalla S, et al. Oncogenic human papillomaviruses block expression of the B-cell translocation gene-2 tumor suppressor gene. Int J Cancer, 2009, 125(9):2014-2020. doi: 10.1002/ijc.24671 [4] Li F, Liu J, Park ES, et al. The B cell translocation gene(BTG) family in the rat ovary:hormonal induction, regulation, and impact on cell cycle kinetics. Endocrinology, 2009, 150(8):3894-3902. doi: 10.1210/en.2008-1650 [5] Zhang YJ, Wei L, Liu M, et al. BTG2 inhibits the proliferation, invasion, and apoptosis of MDA-MB-231 triple-negative breast cancer cells. Tumour Biol, 2013, 34(3):1605-1613. [6] Wagener N, Bulkescher J, Macher-Goeppinger S, et al. Endogenous BTG2 expression stimulates migration of bladder cancer cells and correlates with poor clinical prognosis for bladder cancer patients. Br J Cancer, 2013, 108(4):973-982. [7] Hu XD, Meng QH, Xu JY, et al. BTG2 is an LXXLL-dependent co-repressor for androgen receptor transcriptional activity. Biochem Biophys Res Commun, 2011, 404(4):903-909. doi: 10.1016/j.bbrc.2010.12.064 [8] Fan S, Meng Q, Saha T, et al. Low concentrations of diindolylmethane, a metabolite of indole-3-carbinol, protect against oxidative stress in a BRCA1-dependent manner. Cancer Res, 2009, 69(15):6083-6091. doi: 10.1158/0008-5472.CAN-08-3309 -

 点击查看大图
点击查看大图

图(6)
计量
- 文章访问数: 2003
- HTML全文浏览量: 926
- PDF下载量: 2