


 下载:
下载:


-
肺癌是世界范围内致死率较高的恶性肿瘤之一,近年来肺癌在我国的发病率和病死率均高居首位,其主要原因在于肺癌的高侵袭转移能力和高复发率[1]。非小细胞肺癌是肺癌中发病率最高的类型,手术治疗和放化疗是最常用和最基本的治疗手段,但预后依然较差,5年生存率不足17%[2]。因此,研究肺癌的代谢及转移机制在一定程度上能提高肺癌的诊疗效果,进而降低肺癌的病死率。
在RNA中,大部分为非编码RNA,其中长链非编码RNA(long non-coding RNA,lncRNA)长度一般超过200 bp,且缺乏明显的开放阅读框,因此其不编码蛋白质,但是广泛参与人体生理、病理活动,包括参与或介导肿瘤的发生、发展过程[3-3]。早期研究结果发现,lncRNA尿路上皮癌胚抗原(urothelial carcinoma associated,UCA)1在多种肿瘤细胞中表达上调,降低UCA1的表达可以抑制肿瘤细胞的增殖、转移等生物学行为,调节肿瘤细胞的葡萄糖代谢[4],因此UCA1可作为肿瘤早期诊断、预后监测的新型生物标志物[5-6]。
本研究首先通过检测lncRNA UCA1在肺腺癌A549细胞中的表达,然后体外实验进一步验证lncRNA UCA1在肺腺癌中的作用,旨在进一步揭示肺癌的发生、发展机制,并为lncRNA UCA1作为肺癌新的早期诊断指标及治疗靶点提供证据。
-
肺腺癌A549细胞购自中国科学院上海生命科学研究院,人正常支气管上皮细胞HBE购自中国科学院细胞库,胎牛血清购自美国CLACK公司,DMEM/F12细胞培养基、青-链霉素双抗和胰蛋白酶均购自美国Hyclone公司,Opti-MEM低血清培养基、Lipofectamine 3000和Trizol试剂均购自美国Invitrogen公司,SYBR Premix Ex Taq Ⅱ试剂盒(RR820A)、PrimeScriptTM RT reagent试剂盒(RR047A)购自日本Takara公司,BCA蛋白浓度测定试剂盒、十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)凝胶快速制备试剂盒购自上海碧云天生物技术公司,抗体均购自美国ABcam公司,Transwell小室购自美国Corning公司。18F-FDG由本科室的美国GE公司的MINItrace 加速器合成,放射化学纯度>95%。Image Lab图像分析系统为美国伯乐公司生产,γ计数器(GC-1500)为安徽中科中佳科学仪器有限公司生产。
-
人正常支气管上皮细胞HBE培养于含有10%胎牛血清的1640培养基中,肺腺癌A549细胞培养于含10%胎牛血清和1%双抗DMEM/F12完全培养基中,均置于37℃的恒温培养箱中培养(内含5%CO2);然后加1 mL胰蛋白酶消化收集细胞,再用1 mL无双抗的完全培养基中和,重悬为细胞悬液,按1×105个数量接种于六孔板中,待细胞密度约为70%时,进行转染。将小干扰RNA(small interfering,siRNA)转染细胞分为NC组(阴性对照组,转染siRNA-UCA1序列)和siRNA-UCA1组(转染siRNA-UCA1敲降序列),以上序列由上海吉凯公司设计合成。转染24 h后换液,继续培养24 h后用Trizol试剂裂解细胞,用于检测干扰效率。
-
提取人正常支气管上皮细胞HBE和肺腺癌A549细胞的RNA,进行反转录反应(去除基因组DNA和反转录反应)和实时荧光定量(quantitative real-time,qRT)PCR反应,后者包括预变性95℃ 30 s,反应40个循环,溶解95℃ 5 s、60℃ 30 s、72℃ 5 s、4℃降温30 s等步骤。并以甘油醛-3-磷酸脱氢酶(GAPDH)为内参,通过2−△△CT法计算以上2种细胞的UCA1相对表达量,检测肺腺癌A549细胞的糖代谢相关指标:己糖激酶(hexokinase,HK)2、葡萄糖转运蛋白(glucose transporter protein,GLUT)1和丙酮酸激酶(pyruvate kinase isozymes M,PKM)2的表达量,以β-actin为内参。引物序列如下:
GAPDH引物序列:
正向5′-GAGTCAACGGATTTGGTCGT-3′
反向5′-TTGATTTTGGAGGGATCTCG-3′
β-actin引物序列:
正向5′-GCAGAAGGAGATCACTGCCCT-3′
反向5′-GCTGATCCACATCTGCTGGAA-3′
UCA1引物序列:
正向5′-CTCTCCATTGGGTTCACCATTC-3′
反向5′-GCGGCAGGTCTTAAGAGATGAG-3′
GLUT1引物序列:
正向5′-ATACTCATGACCATCGCGCTAG-3′
反向5′-AAAGAAGGCCACAAAGCCAAAG-3′
HK2引物序列:
正向5′-GGCAATGAAACCAAAGCCAGGAG-3′
反向5′-GGAAGGAGGAGCCAGAAGCAACC-3′
PKM2引物序列:
正向5′-GGGTTCGGAGGTTTGATG-3′
反向5′-ACGGCGGTGGCTTCTGT-3′
-
肺腺癌A549细胞转染48 h后,先用胰蛋白酶消化,再用无血清培养基稀释,以2×105个的数量加入上室中。在下室加入600 μL含20%血清的DMEM/F12培养基,将上室放入后继续培养。8 h后取出上室,用4%的多聚甲醛固定30 min,结晶紫染色30 min,用PBS进行清洗后,擦去上室内残留细胞,晾干拍照。实验至少重复3次。
细胞转染48 h后进行划痕实验,用10 µL加样器的枪头垂直于六孔板划一道竖线,PBS清洗1次后进行拍照,更换无血清培养基培养24 h后再进行拍照,尽量选择同一视野。使用Image J软件(
http://rsb.info.nih.gov/ij/download.html )测各个时间点细胞未覆盖的面积。划痕愈合率(%)=1−(各时间点划痕面积/开始的划痕面积×100%)
-
肺腺癌A549细胞转染48 h后,用PBS清洗再加入裂解液,冰上裂解30 min,4℃离心(17 000×g)25 min后取上清,进行蛋白浓度测定,电泳仪设置电压为80 V跑电泳,至分离胶和浓缩胶交界处时将电压更改为120 V,至样品跑出即可停止电泳。5%脱脂奶封闭2 h,一抗为HK2(1∶1000)、GLUT1(1∶2000)、PKM2(1∶1000),以β-actin(1∶10000)为内参,4℃ 过夜后,第2天用TBST(Tris buffered Tween)缓冲液清洗3次,每次15 min。山羊抗兔二抗(1∶5000)室温孵育2 h,TBST清洗3次后进行化学发光法检测和曝光拍照,结果用image-Lab图像分析系统测定各条带的灰度值,计算HK2、GLUT1和PKM2的相对表达量,实验重复3次。
-
六孔板内未转染的肺腺癌A549细胞每孔加入37 000 Bq的18F-FDG,37℃分别孵育5、15、30、60、90、120 min,计算不同时间点18F-FDG的摄取率,选取摄取率最高的时间点进行后续实验。肺腺癌A549细胞在六孔板内转染48 h后,每孔加入0.296 MBq(8 μCi)/mL的18F-FDG,37℃孵育60 min,吸取上清液后用1 mL的PBS清洗3次。用胰蛋白酶消化细胞,收集后用1 mL的PBS再清洗3次,均加入试管中,最后用γ计数器测量每个试管内的放射性计数,计算细胞对18F-FDG的摄取率:18F-FDG摄取率=细胞内放射性计数/(细胞内放射性计数+上清液放射性计数)×100%。
-
以上所有实验均重复3次以确保结果的可靠性。采用SPSS20.0软件和GraphPad Prism7.0软件对数据进行统计学分析。符合正态分布的计量资料用均数±标准差表示,方差齐的2组数据的比较采用t检验。P<0.05为差异有统计学意义。
-
用qRT-PCR检测人正常支气管上皮细胞HBE和肺腺癌A549细胞中UCA1的表达时发现,UCA1在A549细胞中的表达显著升高,是在HBE中表达量的4.26倍,且差异有统计学意义(t=16.26,P<0.001)。
-
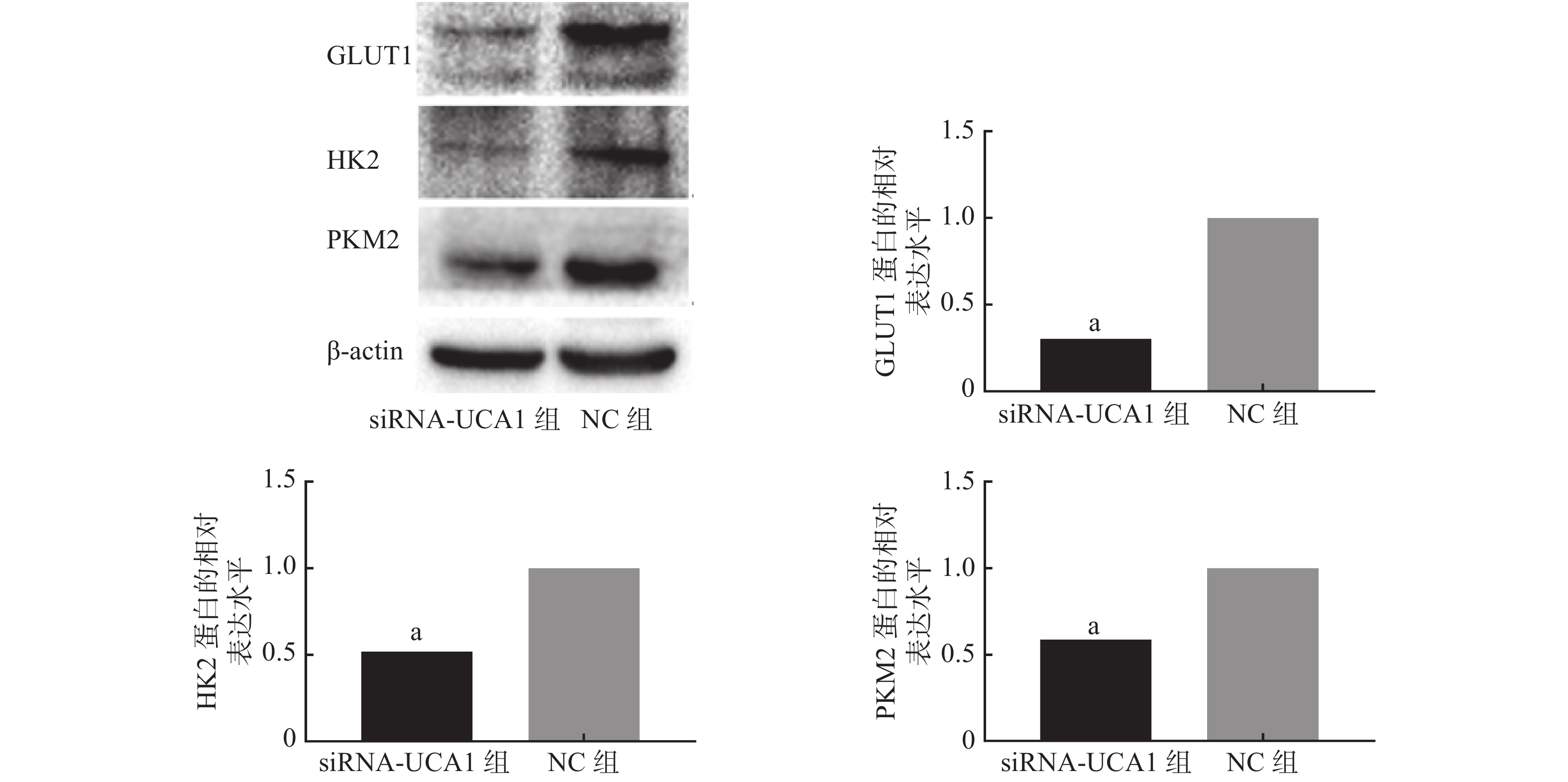
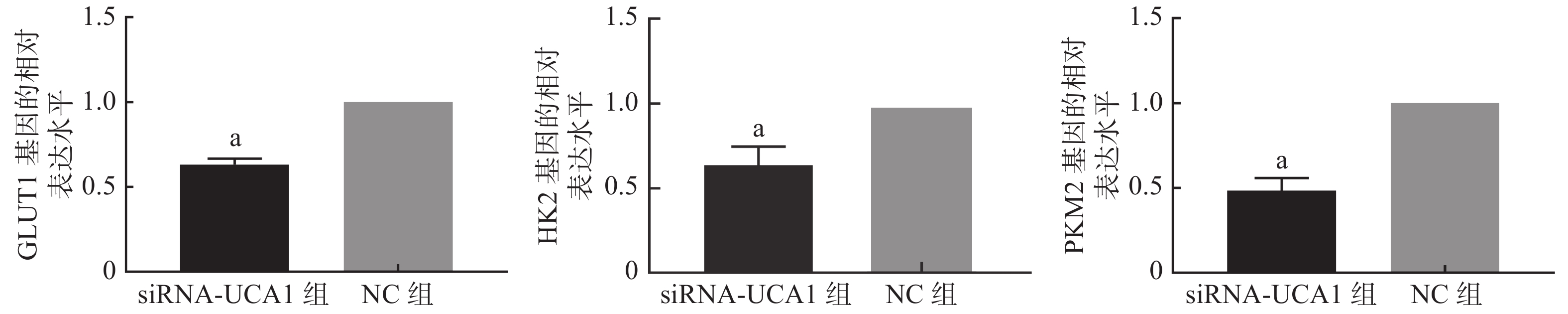
qRT-PCR结果显示,UCA1的siRNA转染能够抑制肺腺癌A549细胞GLUT1、HK2和PKM2基因的表达(图1)。进一步的Western blot结果表明,与NC组相比,siRNA-UCA1组的GLUT1、HK2和PKM2蛋白的表达水平均明显下降,且差异均有统计学意义(图2)。
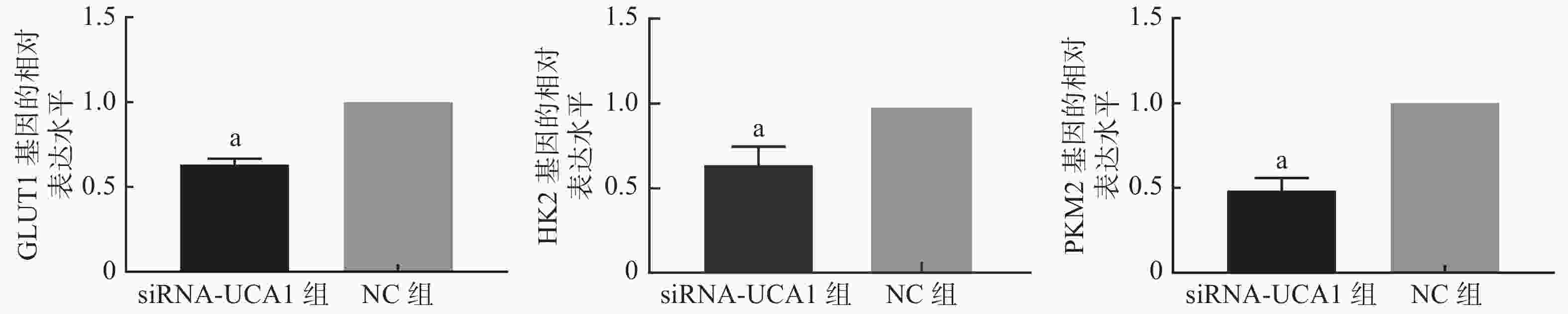
图 1 qRT-PCR检测3种基因的表达情况
Figure 1. qRT-PCR was used to detect the expression of three genes
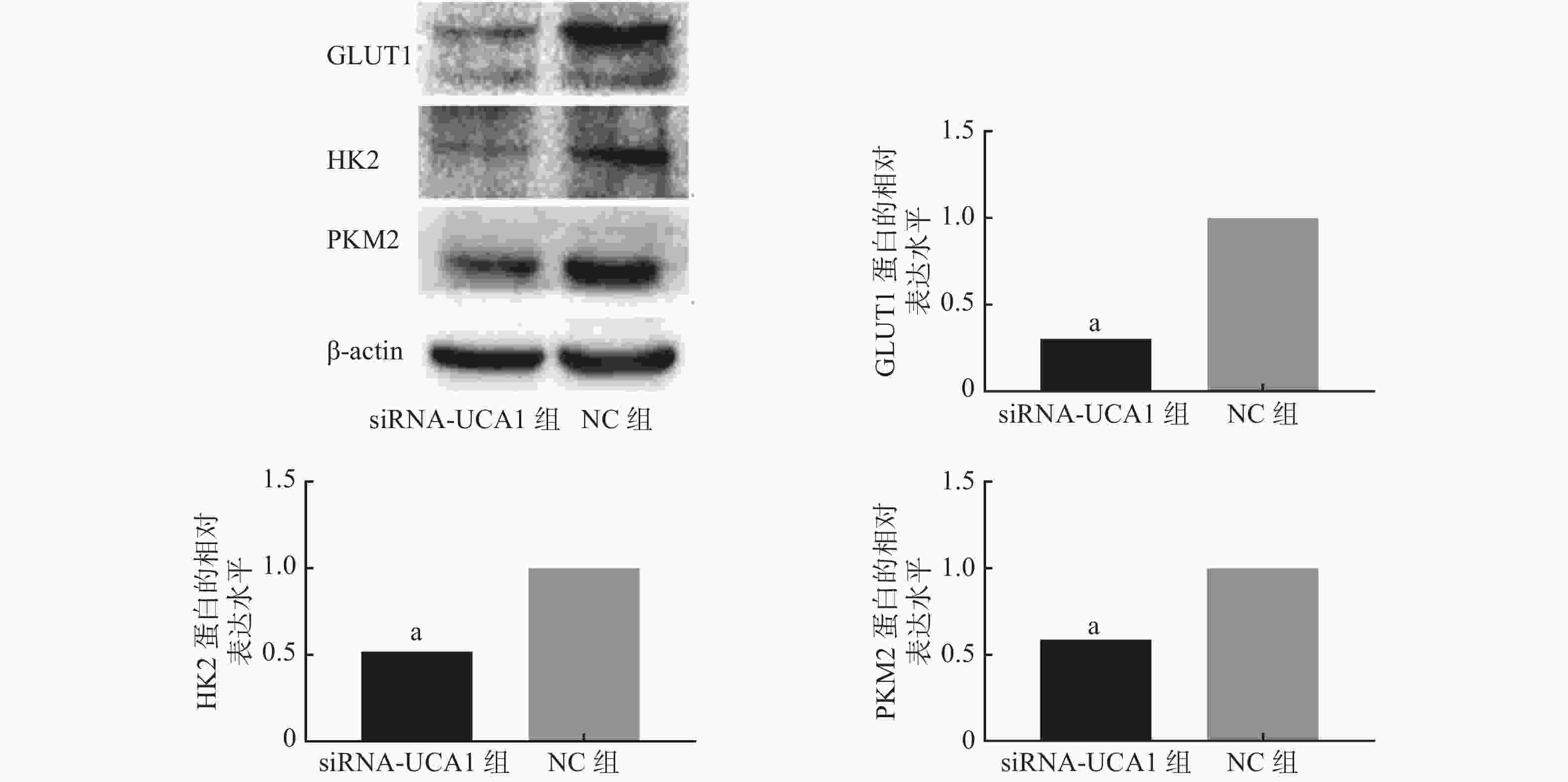
图 2 Western blot检测3种蛋白的表达情况
Figure 2. Western blot was used to detect the expression of three proteins
-
Transwell侵袭实验结果显示,siRNA-UCA1组和NC组细胞穿过数量分别为(12.0000±0.5774)、(127.7000±5.9250)个(图3),说明下调UCA1时,肺腺癌细胞的侵袭能力与对照组相比显著降低(t=19.43,P<0.001)。
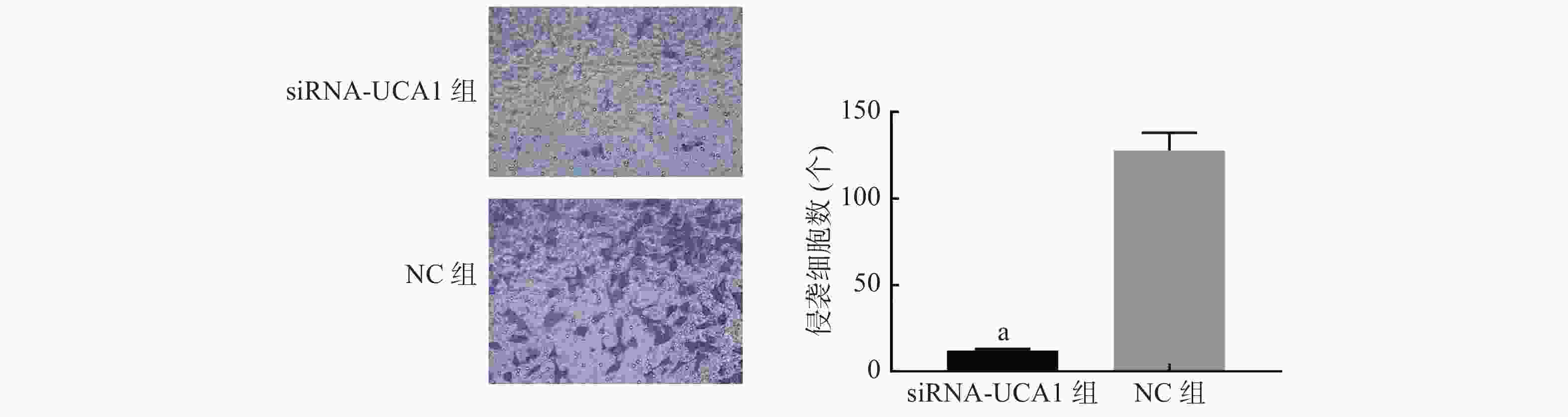
图 3 siRNA-UCA1对肺腺癌A549细胞侵袭能力的影响(结晶紫染色,×400)
Figure 3. Effect of siRNA-urothelial carcinoma associated 1 on the invasive ability of lung adenocarcinoma A549 cells(crystal violet staining, ×400)
-
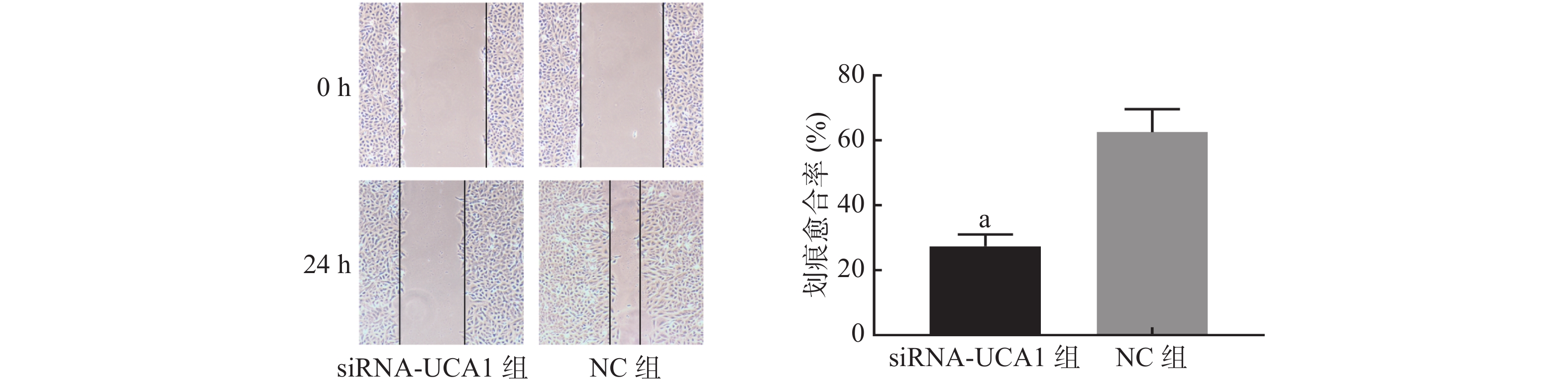
划痕实验结果显示,经过24 h的爬行,与NC组相比,siRNA-UCA1组细胞的迁移运动能力明显降低(图4)。
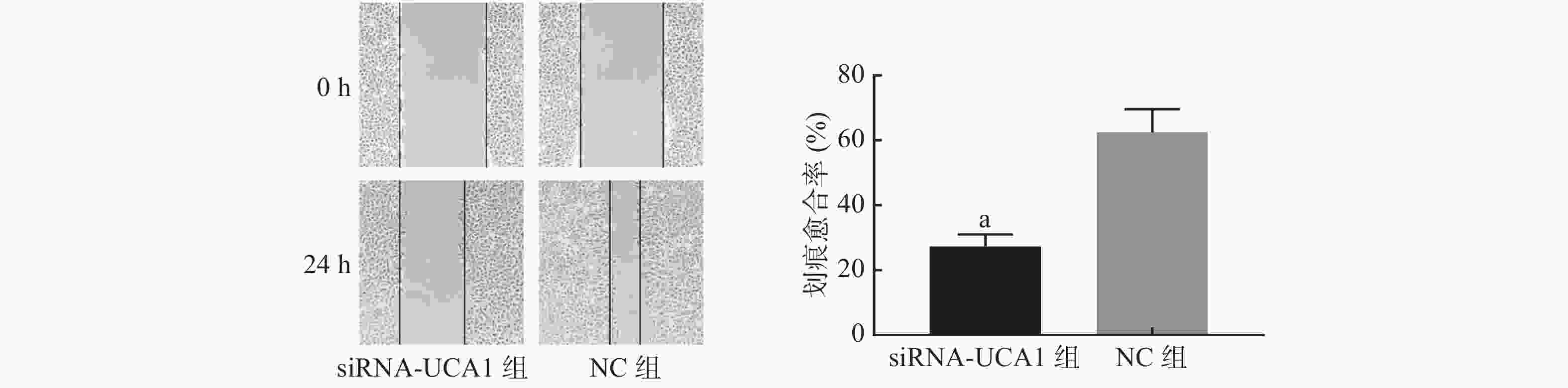
图 4 siRNA-UCA1对肺腺癌A549细胞迁移能力的影响(×100)
Figure 4. Effect of siRNA-urothelial carcinoma associated 1 on the migration ability of lung adenocarcinoma A549 cells(×100)
-
未转染的肺腺癌A549细胞在5、15、30、60、90、120 min时对18F-FDG的摄取率分别为1.95%、2.28%、2.55%、2.86%、2.40%、2.34%,60 min时的摄取率达到峰值,因此选择60 min作为测定的时间点。转染后的肺腺癌A549细胞测定结果如图5显示,siRNA-UCA1组对18F-FDG的摄取率较NC组明显降低,且差异有统计学意义。
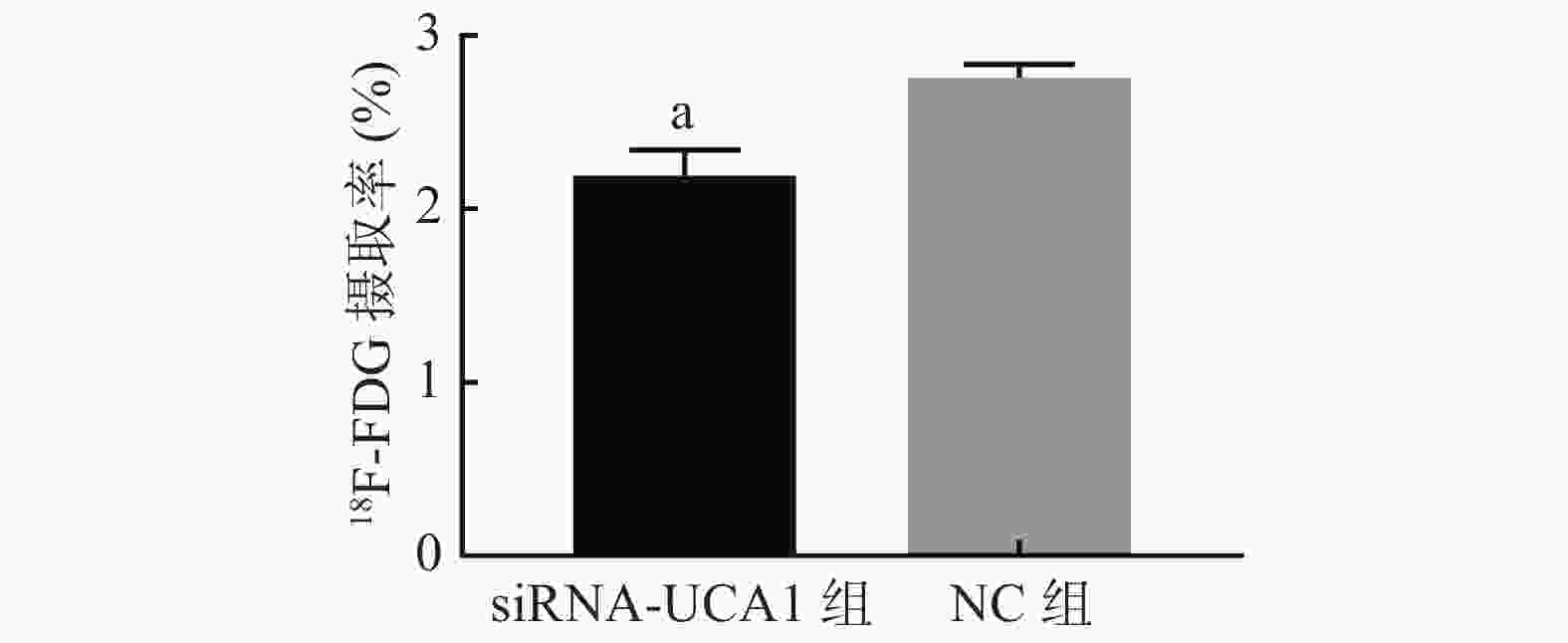
图 5 siRNA-UCA1对肺腺癌A549细胞18F-FDG摄取率的影响
Figure 5. Effect of siRNA-urothelial carcinoma associated 1 on the 18F-FDG uptake rate of lung adenocarcinoma A549 cells
-
目前针对恶性肿瘤的治疗手段包括手术切除并行同步放化疗,但整体预后仍不理想[7]。因此,研究肺癌病变的确切机制和发现新的关键靶点至关重要,对开展肺腺癌患者的个体化治疗和改善预后具有重要意义。
代谢重编程是肿瘤细胞的标志之一,并且与细胞生长调节的过程密切相关[8]。FDG是一类葡萄糖类似物,可以反映葡萄糖的代谢情况。近年来随着PET类的医学成像设备被临床广泛应用,FDG可以用于肿瘤的诊断、分期和治疗监测。由于细胞在快速增殖侵袭过程中所需的能量会急剧增加,可加快肿瘤发生发展的进程[9-10],因此研究肿瘤的代谢调节过程有利于寻找更好的治疗靶点。而lncRNA最开始被认为是基因转录的副产物,不存在生物学功能[11]。但近年来的研究结果表明,lncRNA与肿瘤的发生发展有密切关系,已成为基因组学新的研究热点[12-13]。
目前,有研究结果表明UCA1在肺癌细胞中呈高表达状态,并且可以促进小细胞肺癌的生长、侵袭和迁移[14-15],与本研究的结果一致。除此之外,本研究还通过qRT-PCR及Western blot实验检测UCA1在A549细胞糖代谢中的关键酶,发现下调UCA1后,GLUT1、HK2和PKM2表达均显著降低,抑制细胞糖代谢的过程,这在国内外尚无报道。我们用Transwell侵袭实验和划痕实验进行生物学行为的检测,结果表明,下调UCA1后会使A549细胞的侵袭和迁移能力显著下降,但具体机制仍需要进一步探索。有研究者发现,UCA1在胰腺癌中可以通过Hippo通路促进胰腺癌细胞侵袭和转移[16];对肝细胞癌的研究发现,UCA1可以通过与miR-203结合,激活转录因子Snail 2,促进肝癌细胞的增殖和侵袭[17]。因此,UCA1沉默后,相应糖代谢指标表达降低,抑制肿瘤的能量代谢,使肿瘤侵袭转移能力下降。
综上所述,本研究发现UCA1可以影响糖代谢的关键酶,并且促进肺腺癌A549细胞的迁移和侵袭,这提示其可能在肺癌发生过程中起到癌基因的作用,有望成为肺癌治疗的潜在新靶点。本研究结果为进一步深入探讨UCA1对促进肺癌细胞恶性生物学特征的分子机制奠定了基础。由于我们只进行了体外实验,尚存在不足之处,比如UCA1与临床病理因素及预后的关系等都值得进一步研究。接下来将进行机制研究,深入探讨UCA1通过哪种信号通路对糖代谢指标进行调节,同时加上体外动物实验进行验证,以得到更明确的治疗效果。
利益冲突 本研究由署名作者按以下贡献声明独立开展,不涉及任何利益冲突。
作者贡献声明 张卉负责试验的设计与实施、论文的撰写;王姝负责数据的分析;李雪娜、李亚明负责论文的审阅与修订。
长链非编码RNA UCA1对肺腺癌A549细胞糖代谢及侵袭转移的影响
The functional role of long non-coding RNA UCA1 in lung adenocarcinoma A549 cells on glucose metabolism, invasion and metastasis
-
摘要:
目的 探讨长链非编码RNA(lncRNA)尿路上皮癌胚抗原1(UCA1)在肺腺癌A549细胞中的表达及对糖代谢关键酶和细胞侵袭、转移的影响。 方法 构建lncRNA UCA1沉默及其对照稳定转染的肺腺癌A549细胞系,将小干扰RNA(siRNA)转染细胞分为NC组(阴性对照组,转染siRNA-UCA1序列)和siRNA-UCA1组(转染siRNA-UCA1敲降序列)。采用实时荧光定量PCR、Western blot检测糖代谢相关指标葡萄糖转运蛋白(GLUT)1、己糖激酶(HK)2和丙酮酸激酶(PKM)2水平的变化。采用Transwell实验和划痕实验检测siRNA-UCA1的侵袭和迁移能力。采用18F-氟脱氧葡萄糖(FDG)摄取实验检测siRNA-UCA1的摄取率。计量资料的比较采用t检验。 结果 UCA1在肺腺癌A549细胞中高表达。与NC组相比,siRNA-UCA1组能够抑制GLUT1、HK2和PKM2基因在肺腺癌A549细胞中的表达(t=19.66、5.81、11.98,均P<0.001),且三者的蛋白表达水平明显下降(t=61.35、145.90、88.19,均P<0.001)。与NC组相比,siRNA-UCA1组肺腺癌A549细胞的侵袭能力、迁移运动能力和对18F-FDG的摄取率均明显降低(t=19.43、7.71、5.79,均P<0.05)。 结论 LncRNA UCA1能够抑制糖代谢的关键酶并促进肺腺癌的转移能力,其可能成为肺癌新的诊断指标和治疗靶点。 Abstract:Objective To explore the expression of long non-coding RNA (lncRNA) urothelial carcinoembryonic antigen 1 (UCA1) in lung adenocarcinoma A549 cells and its influence on key enzymes involved in sugar metabolism, cell invasion, and metastasis. Methods The lncRNA UCA1 silencing and its control stable transfected lung adenocarcinoma A549 cells were constructed, the small interfering RNA (siRNA) transfected cells were divided into NC group (negative control group, transfected with siRNA-UCA1 sequence) and siRNA-UCA1 group (transfected with siRNA-UCA1 knockdown sequence). Quantitative real-time PCR and Western blot analyses were used to detect changes in the levels of glucose transporter protein (GLUT)1, hexokinase (HK)2 and pyruvate kinase isozymes M (PKM)2 related to glucose metabolism. Transwell and scratch tests were conducted to detect the invasion and migration ability of siRNA-UCA1. 18F-Fluorodeoxyglucose (FDG) uptake experiment was used to detect the uptake rate of siRNA-UCA1. T test was used to compare the measurement data. Results UCA1 was highly expressed in lung adenocarcinoma A549 cell line. Compared with the NC group, the siRNA-UCA1 group can inhibit the expression of GLUT1, HK2, and PKM2 in lung adenocarcinoma A549 cells (t=19.66, 5.81, 11.98; all P<0.001), and the protein expression levels of the three significantly decreased (t=61.35, 145.90, 88.19; all P<0.001). Compared with the NC group, the siRNA-UCA1 group's migration ability and 18F-FDG uptake rate of A549 cells were significantly reduced (t=19.43, 7.71, 5.79; all P<0.05). Conclusions LncRNA UCA1 can affected the key enzymes of glucose metabolism and promoted the metastasis ability of lung adenocarcinoma. Thus, lncRNA UCA1 may be a new diagnostic index and therapeutic target for lung cancer. -
图 1 qRT-PCR检测3种基因的表达情况
Figure 1. qRT-PCR was used to detect the expression of three genes
图 2 Western blot检测3种蛋白的表达情况
Figure 2. Western blot was used to detect the expression of three proteins
图 3 siRNA-UCA1对肺腺癌A549细胞侵袭能力的影响(结晶紫染色,×400)
Figure 3. Effect of siRNA-urothelial carcinoma associated 1 on the invasive ability of lung adenocarcinoma A549 cells(crystal violet staining, ×400)
图 4 siRNA-UCA1对肺腺癌A549细胞迁移能力的影响(×100)
Figure 4. Effect of siRNA-urothelial carcinoma associated 1 on the migration ability of lung adenocarcinoma A549 cells(×100)
-
[1] Chen WQ, Zheng RS, Baade PD, et al. Cancer statistics in China, 2015[J]. CA Cancer J Clin, 2016, 66(2): 115−132. DOI: 10.3322/caac.21338. [2] Grose D, Devereux G, Milroy R. Comorbidity in lung cancer: important but neglected. a review of the current literature[J]. Clin Lung Cancer, 2011, 12(4): 207−211. DOI: 10.1016/j.cllc.2011.03.020. [3] Peschansky VJ, Wahlestedt C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation[J]. Epigenetics, 2014, 9(1): 3−12. DOI: 10.4161/epi.27473. [4] Xiao ZD, Zhuang L, Gan B. Long non-coding RNAs in cancer metabolism[J]. Bioessays, 2016, 38(10): 991−996. DOI: 10.1002/bies.201600110. [5] Li ZK, Li X, Wu SZ, et al. Long non-coding RNA UCA1 promotes glycolysis by upregulating hexokinase 2 through the mTOR-STAT3/microRNA143 pathway[J]. Cancer Sci, 2014, 105(8): 951−955. DOI: 10.1111/cas.12461. [6] Nie W, Ge HJ, Yang XQ, et al. LncRNA-UCA1 exerts oncogenic functions in non-small cell lung cancer by targeting miR-193a-3p[J]. Cancer Lett, 2016, 371(1): 99−106. DOI: 10.1016/j.canlet.2015.11.024. [7] Nieder C, Guckenberger M, Gaspar LE, et al. Management of patients with brain metastases from non-small cell lung cancer and adverse prognostic features: multi-national radiation treatment recommendations are heterogeneous[J/OL]. Radiat Oncol, 2019, 14(1): 33[2019-09-01]. https://ro-journal. biomedcentral.com. DOI: 10.1186/s13014-019-1237-9. [8] Hanahan D, Weinberg RA. Hallmarks of cancer: The next generation[J]. Cell, 2011, 144(5): 646−674. DOI: 10.1016/j.cell.2011.02.013. [9] Miller DM, Thomas SD, Islam A, et al. c-Myc and cancer metabolism[J]. Clin Cancer Res, 2012, 18(20): 5546−5553. DOI: 10.1158/1078-0432.CCR-12-0977. [10] Zhang J, Goliwas KF, Wang WJ, et al. Energetic regulation of coordinated leader-follower dynamics during collective invasion of breast cancer cells[J]. Proc Natl Acad Sci USA, 2019, 116(16): 7867−7872. DOI: 10.1073/pnas.1809964116. [11] Ricciuti B, Mencaroni C, Paglialunga L, et al. Long noncoding RNAs: new insights into non-small cell lung cancer biology, diagnosis and therapy[J]. Med Oncol, 2016, 33(2): 18. DOI: 10.1007/s12032-016-0731-2. [12] Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs[J]. Cell, 2009, 136(4): 629−641. DOI: 10.1016/j.cell.2009.02.006. [13] Kretz M, Siprashvili Z, Chu C, et al. Control of somatic tissue differentiation by the long non-coding RNA TINCR[J]. Nature, 2013, 493(7431): 231−235. DOI: 10.1038/nature11661. [14] 蔡华荣, 王志强, 江跃全. 长链非编码RNA UCA1靶向调控miR-185-5p对非小细胞肺癌A549细胞的作用及其机制[J]. 中国肿瘤生物治疗杂志, 2018, 25(6): 555−561. DOI: 10.3872/j.issn.1007-385X.2018.06.002.
Cai HR, Wang ZQ, Jiang YQ. Effect and mechanism of long non-coding RNA UCA1 targeting mir-185-5p on non-small cell lung cancer A549 cells[J]. Chin J Cancer Biother, 2018, 25(6): 555−561. DOI: 10.3872/j.issn.1007-385X.2018.06.002.[15] 姜文瑞, 杨方, 吴运福. 长链非编码RNA UCA1介导调控Wntβ-Catenin信号通路对 NSCLS增殖、凋亡及侵袭性的研究[J]. 医学分子生物学杂志, 2019, 16(4): 303−309. DOI: 10.3870/j.issn.1672-8009.2019.04.002.
Jiang WR, Yang F, Wu YF. Effect of long-chain non-coding RNA UCAl targeting regulation of WntB-catenin signaling pathway on proliferation, apoptosis and invasiveness of NSCLS[J]. J Med Mol Biol, 2019, 16(4): 303−309. DOI: 10.3870/j.issn.1672-8009.2019.04.002.[16] Zhang MT, Zhao Y, Zhang YL, et al. LncRNA UCA1 promotes migration and invasion in pancreatic cancer cells via the Hippo pathway[J]. Biochim Biophys Acta Mol Basis Dis, 2018, 1864(5): 1770−1782. DOI: 10.1016/j.bbadis.2018.03.005. [17] Xiao JN, Yan TH, Yu RM, et al. Long non-coding RNA UCA1 regulates the expression of Snail2 by miR-203 to promote hepatocellular carcinoma progression[J]. J Cancer Res Clin Oncol, 2017, 143(6): 981−990. DOI: 10.1007/s00432-017-2370-1. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3650
- HTML全文浏览量: 2354
- PDF下载量: 8