



-
结直肠癌(colorectal cancer,CRC)是一种发病率和致死率较高的常见消化道恶性肿瘤,呈逐年上升趋势,其发病率在全世界范围内已位居第2位、致死率已位居第5位[1]。CRC发病与年龄、性别、遗传、结直肠息肉等多种因素相关,主要通过结肠镜和粪便潜血实验进行筛查[2-3]。放疗(包括新辅助放疗)是CRC临床治疗的重要手段,但是部分CRC患者对放疗不敏感甚至存在放疗抵抗,是影响CRC患者预后和生活质量的重要原因[4]。
核仁小RNA(small nucleolar RNA,snoRNA)是一类主要存在于细胞核仁中的小分子非编码RNA,大小约为60~300 个核苷酸,其主要生物学功能为调节核糖体生成加工过程[5]。snoRNA在多种癌症中发挥关键作用,比如SNORA21、SNORA42、SNORA71A等snoRNA在CRC中高表达,通过调节细胞增殖、凋亡、侵袭和迁移等过程发挥促癌作用[6–9],但是snoRNA在肿瘤放射敏感性中的作用仍不清楚。SNORA72(基因ID: ENSG00000207067)是由RPL30基因编码、长度为132 bp的snoRNA分子,在人类、恒河猴、小鼠、犬和大象等基因组中具有较高的同源性。Zhang等[10]的研究结果显示,SNORA72能促进卵巢癌细胞的干性转化,但是其在CRC发生发展以及放射敏感性中的作用未见报道。
本研究通过体外细胞实验分析SNORA72对CRC细胞增殖、存活及放射敏感性的影响,旨在探索SNORA72在CRC组织中的表达水平及其与患者预后的相关性。
-
DMEM培养基购自德国Sigma公司;胎牛血清购自上海ExCellBio公司;反义寡核苷酸(antisense oligonucleotide,ASO)及细胞转染试剂riboFECT™CP购自广州市锐博生物科技有限公司;慢病毒颗粒购自上海吉凯公司;Trizol试剂购自德国Sigma公司;反转录试剂盒PrimeScript RT reagent kit with gDNA Eraser购自日本Takara公司;荧光定量PCR试剂盒2×M5 HIPer SYBR Premix EsTaq购自北京聚合美生物科技有限公司;碘化丙啶(propidiumiodide,PI)染液购自天津三箭生物技术有限公司;膜联蛋白V(Annexin V)-异硫氰酸荧光素(fluorescein isothiocyanate,FITC)/碘化丙啶(propidium iodide,PI)双染法细胞凋亡检测试剂盒购自日本Dojindo公司;Cell Counting Kit-8细胞增殖检测试剂盒购自日本Dojindo公司;BCA(bicinchoninic acid)蛋白定量检测试剂盒购自碧云天生物技术公司;兔抗人PARP抗体和兔抗人Caspase3抗体购自美国Cell Signaling Technology公司;鼠抗人Bax抗体和鼠抗人Survivine抗体购自美国SantaCruz公司;兔抗人Bcl-2抗体购自美国ImmunoWay公司;鼠抗人β-actin抗体购自美国Proteintech公司;羊抗鼠辣根过氧化物酶标记IgG抗体和羊抗兔辣根过氧化物酶标记IgG抗体购自美国KPL公司;MODEL2300型恒温培养箱购自日本松下电器公司;NovoCyte 2060R型流式细胞仪购自美国艾森生物公司;ImageQuant AI800型超敏化学发光成像仪购自美国 GE 公司。60Co γ 射线源由中国人民解放军军事科学院军事医学研究院辐射医学研究所提供。
-
CRC细胞株HT29购自上海吉凯基因医学科技有限公司。细胞培养条件:用含10%胎牛血清和1%青霉素-链霉素的DMEM培养基在37℃、含5% CO2的恒温培养箱内培养。
-
应用TIMER 2.0数据库(
http://timer.cistrome.org )分析SNORA72在不同癌症组织中的表达变化;通过cBioPortal 网站(http://www.cbioportal.org ) 评估泛癌基因组中SNORA72拷贝数变异;应用GEO数据库(https://www.ncbi.nlm.nih.gov/geo )分析CRC组织和癌旁组织中SNORA72的表达差异;应用TCGA数据库(https://xenabrowser.net/datapages )对SNORA72表达水平和CRC患者临床组织病理学特征进行相关性分析。 -
将CRC细胞株HT29按2.5×105 个/孔接种于6孔板,培养24 h后,用50 μmol/L浓度靶向SNORA72的ASO序列(ASO-SNORA72,干扰序列:5'-CTCGCTGTTCTGATTTTGTA-3')及阴性对照序列(ASO-NC,广州锐博生物技术有限公司提供,货号:lnc6N0000002-1-10),按riboFECT™CP说明书进行转染,转染48 h后收集细胞用于后续实验。将 HT29细胞分为2组:ASO-NC组(表示敲低SNORA72的阴性对照组)和ASO-SNORA72组(表示敲低 SNORA72的表达组)。
-
将CRC细胞株HT29按2.5×105 个/孔接种于6孔板,使用慢病毒颗粒(感染复数=10)的SNORA72过表达病毒(LV-SNORA72,插入基因序列:5'-CTGCGAATATTCTCGCTGTTCTGATTTTGTAATAGTCAGGACAGGCTAAACATTCGCTATATTAAGACCATGCATGTGTCCCCAAACCTAGTTCTTTCCCTAGGTCTGGTTTCATAAATGCTGGTGATAAACTTTTTT-3')及阴性对照病毒(LV-NC,插入基因序列:5'-TTCTCCGAACGTGTCACGT-3')分别感染HT29细胞,于感染后72 h用含2 μg/ml嘌呤霉素的培养基进行阳性细胞筛选,持续筛选2~3 d获得稳定细胞系,并用含0.67 μg/ml嘌呤霉素的培养基维持培养。将 HT29细胞分为 2 组:LV-NC组(表示过表达SNORA72的阴性对照组)和LV-SNORA72组(表示过表达 SNORA72组)。
-
使用60Co γ射线照射源对LV-SNORA72组和LV-NC组的HT29细胞进行照射,剂量率为65.69 cGy/min。
-
使用Trizol试剂提取各组细胞中总RNA,用反转录试剂盒PrimeScript RT reagent kit with gDNA Eraser将RNA反转录成cDNA。采用qRT-PCR试剂盒2×M5 HIPer SYBR Premix EsTaq进行qRT-PCR反应,检测细胞中SNORA72的表达水平。SNORA72引物序列:正向5'-TGCGAATATTCTCGCTGTTCTG-3';反向5'-CGAATGTTTAGCCTGTCCTGAC-3'。U6(内参基因)引物序列:正向5'-CTCGCTTCGGCAGCACA-3';反向5'-AACGCTTCACGAATTTGCGT-3'。通过2−ΔΔCT法计算SNORA72的相对表达量。
-
HT29细胞经0.25%胰酶消化后,900×g 离心5 min后收集细胞,用PBS重悬细胞1次,用70%乙醇于−20℃固定24 h,经900×g 离心5 min后弃固定液,用PBS将细胞漂洗2次后,加200 μl PI染液于37℃避光染色30 min,用流式细胞仪检测细胞周期。
-
在检测敲低SNORA72能否诱导细胞凋亡的实验中,将ASO-NC组和ASO-SNORA72组的HT29细胞经0.25%胰酶消化后,900×g 离心5 min,用PBS漂洗1次,使用Annexin V-FITC/PI双染法细胞凋亡检测试剂盒进行凋亡细胞染色,轻轻混匀后室温避光染色15 min,1 h之内用流式细胞仪检测细胞凋亡情况。在放射诱导细胞凋亡实验中,对LV-NC组和LV-SNORA72组的HT29细胞分别进行0 Gy(假照射)和4、8、12 Gy γ射线单次剂量照射,于照射后48、72 h收获细胞,重复上述步骤以检测细胞凋亡。
-
将对数生长期的细胞以2000 个/孔密度接种于96孔板,每组设5个重复孔,按照Cell Counting Kit-8试剂盒说明书操作,于接种后第0、1、2、3、4、5天分别测定450 nm波长处吸光度值,并绘制细胞增殖曲线。
-
将HT29细胞按一定数量(800~1200个/孔)接种于6孔板,每组设3个重复孔,连续培养10~14 d至细胞克隆形成,用无水甲醇固定细胞并用吉姆萨染液进行染色,在显微镜下统计分析,以细胞数≥50个为1个有效克隆。细胞克隆形成率=(细胞克隆数/初始接种细胞总数)×100%。在放射敏感性分析实验中,于细胞接种后第2天进行0、1、2、4、6 Gy γ射线单次剂量照射,待观察到明显的细胞克隆形成时,用无水甲醇固定、吉姆萨染液染色。计算细胞克隆形成率,通过单击多靶模型拟合细胞存活曲线,计算放射敏感性的相关参数。
-
采用放射免疫沉淀法(radio immunoprecipitation assay,RIPA)裂解缓冲液从转染ASO-SNORA72组及ASO-NC组的HT29细胞中提取全蛋白。采用湿转法90 V恒压转膜完成Western blot实验,用5%脱脂牛奶室温封闭2 h,依次加入兔抗人PARP抗体、兔抗人caspase3抗体、兔抗人Bcl-2抗体、鼠抗人Bax抗体、鼠抗人survivine抗体(均为1∶1000稀释)、鼠抗人β-actin抗体(1∶5000稀释),4℃孵育过夜,用TBST缓冲液洗膜3次,每次5 min;羊抗鼠辣根过氧化物酶标记IgG抗体、羊抗兔辣根过氧化物酶标记IgG抗体(均为1∶5000稀释)室温孵育2 h,TBST缓冲液洗膜3次,每次5 min,经抗体孵育后,用增强型化学发光试剂对目的蛋白条带进行显影,通过超敏化学发光成像仪自动曝光记录结果。
-
收集LV-NC组和LV-SNORA72组的HT29细胞,每组均设3个重复样本,委托杭州联川生物技术公司经IlluminaX10测序平台进行转录组测序。差异基因阈值设置:P<0.05,log2(变化倍数)>1或log2(变化倍数)<−1。应用DAVID网站(
https://david.ncifcrf.gov )对差异基因进行功能及通路富集分析;应用STRING网站(https://string-db.org )对差异基因进行蛋白相互作用分析。 -
应用SPSS 26.0和Graphpad Prism 9.0软件进行统计学分析。数据在进行正态性检验及方差齐性检验后,符合正态分布的数据以
$\bar x$ ±s表示,两组间的比较采用独立样本t检验(方差齐)。SNORA72表达与CRC临床病理特征之间的相关性采用χ2检验,采用Kaplan-Meier法计算总生存率并绘制生存曲线。P<0.05为差异有统计学意义。 -
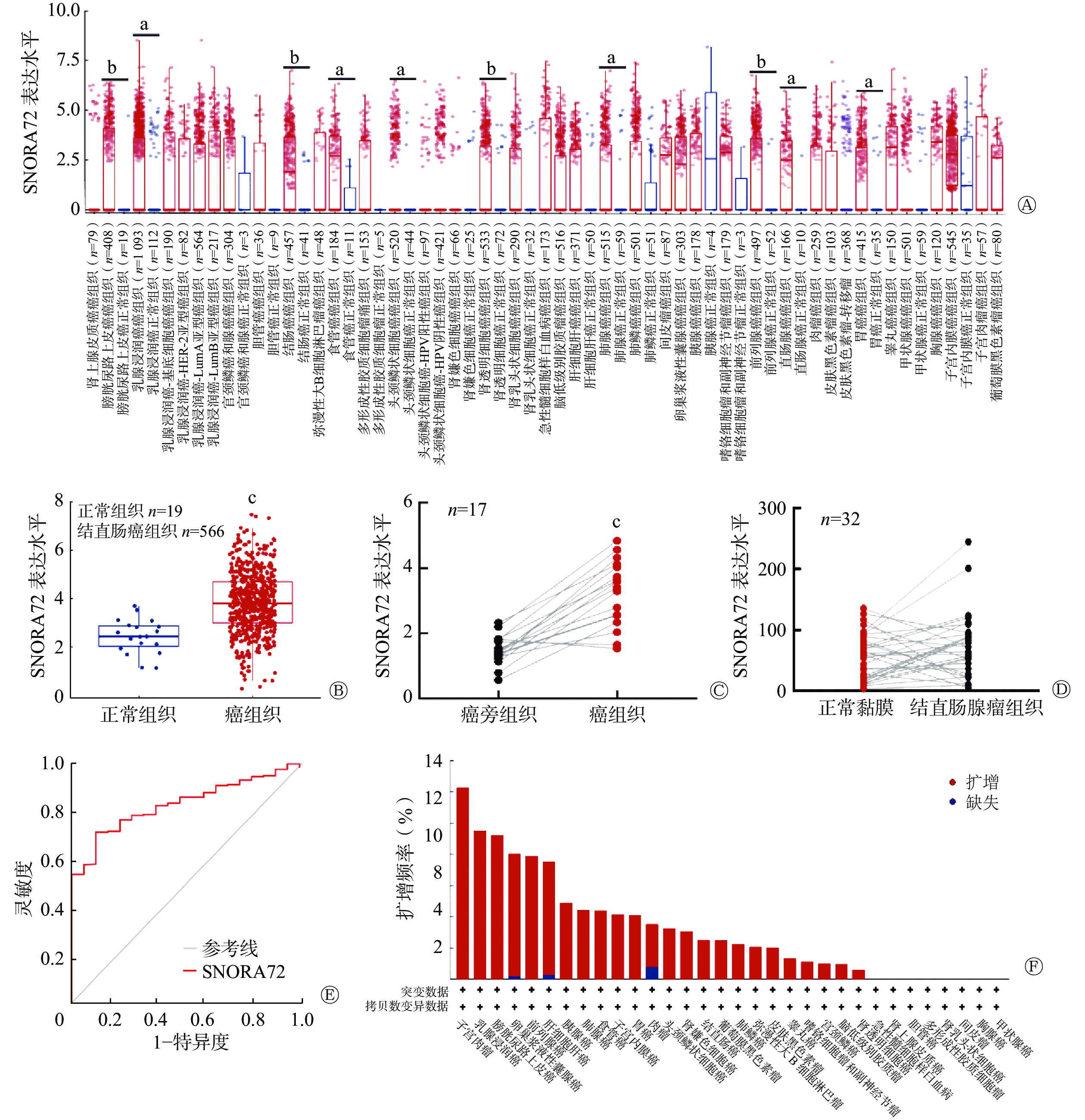
应用TIMER 2.0数据库分析癌和正常组织中 SNORA72 的表达,结果显示 SNORA72 在 10种(膀胱尿路上皮癌、乳腺浸润癌、结肠癌、食管癌、头颈鳞状细胞癌、肾透明细胞癌、肺腺癌、前列腺癌、直肠腺癌和胃癌)癌症类型中上调,且差异均有统计学意义(均P<0.05,图1A)。为进一步分析SNORA72在CRC中的表达,对GEO数据库数据集GSE39582进行分析,该数据集包含了19例正常组织样本和566例CRC组织样本,结果显示,与正常组织相比,SNORA72在CRC组织中的表达水平升高,且差异有统计学意义(t=4.655,P<0.001,图1B)。并且,应用GEO数据库中的GSE32323数据集,对17例CRC患者的癌组织与癌旁组织的SNORA72表达水平进行了配对样本分析,结果显示SNORA72在癌组织中的表达水平高于癌旁组织,且差异有统计学意义(t=7.382,P<0.001,图1C)。此外,应用GSE8671数据集对32例结直肠腺瘤患者的正常黏膜与结直肠腺瘤中SNORA72的表达水平进行配对样本分析,结果显示,SNORA72在正常黏膜与结直肠腺瘤组织中的表达差异无统计学意义(t=1.556,P=0.129,图1D)。以上结果表明,SNORA72在CRC组织中高表达。同时,对GSE39582数据集进行ROC曲线分析,结果显示SNORA72的表达水平能特异性区分癌组织与癌旁组织,AUC为0.837(95% CI:0.777~0.892,P<0.001,图1E)。由于基因组扩增引起的基因拷贝数变异是癌症相关基因异常表达的重要原因[11],应用cBioPortal 数据库对不同癌症基因组中SNORA72拷贝数变异情况进行分析(数据来自4项研究,包括1616例患者),结果显示SNORA72在子宫颈肉瘤、乳腺浸润癌、膀胱尿路上皮癌、卵巢浆液性囊腺癌等24种癌症基因组中普遍扩增(图1F),其中在CRC组织中的扩增频率为2.5%。上述结果提示,SNORA72在CRC等癌组织中高表达可能由基因扩增所致,且其异常高表达对CRC具有一定的分类诊断价值。
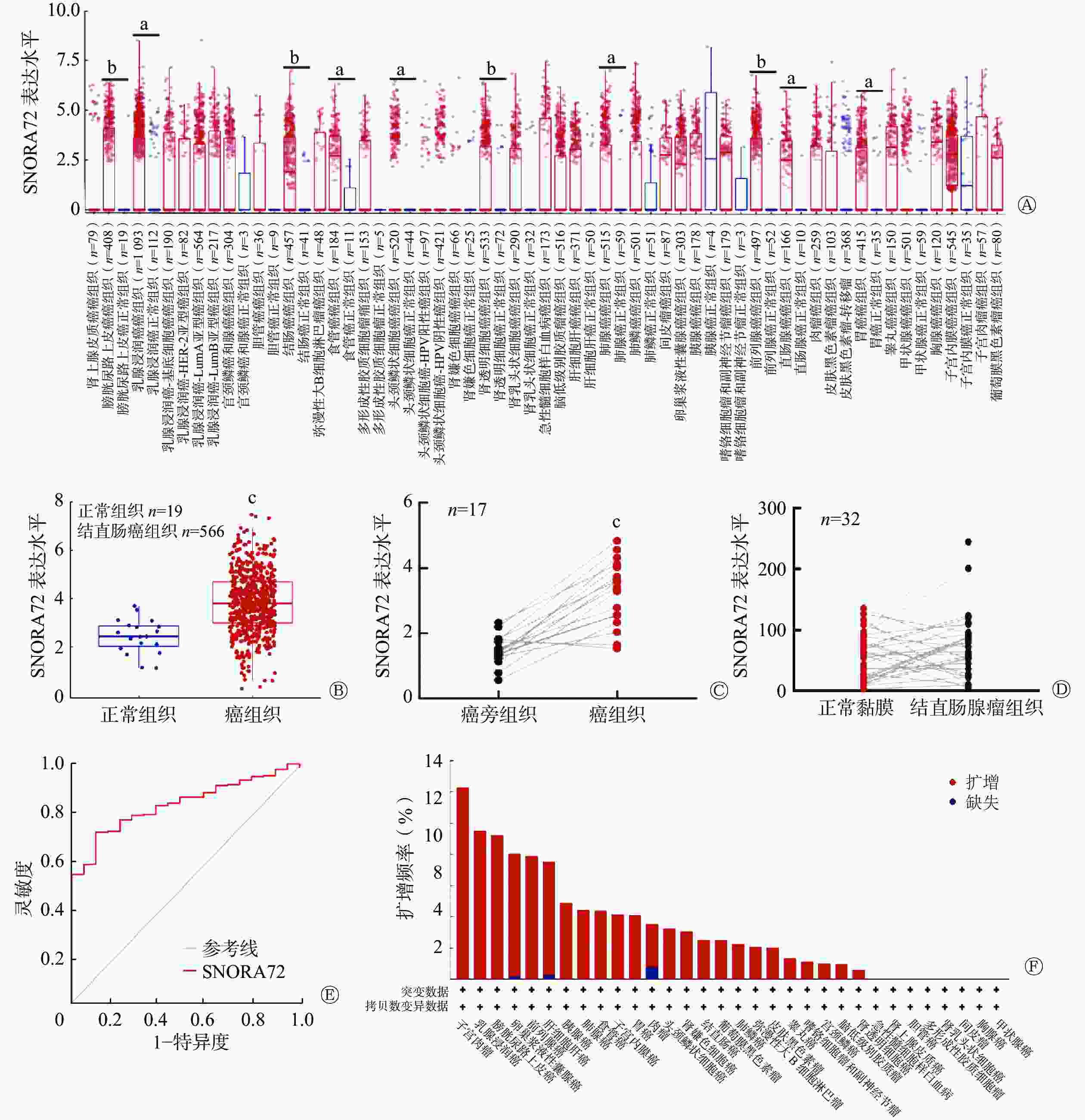
图 1 SNORA72在多种癌组织中的表达情况
Figure 1. Expression of SNORA72 in various cancer tissues
-
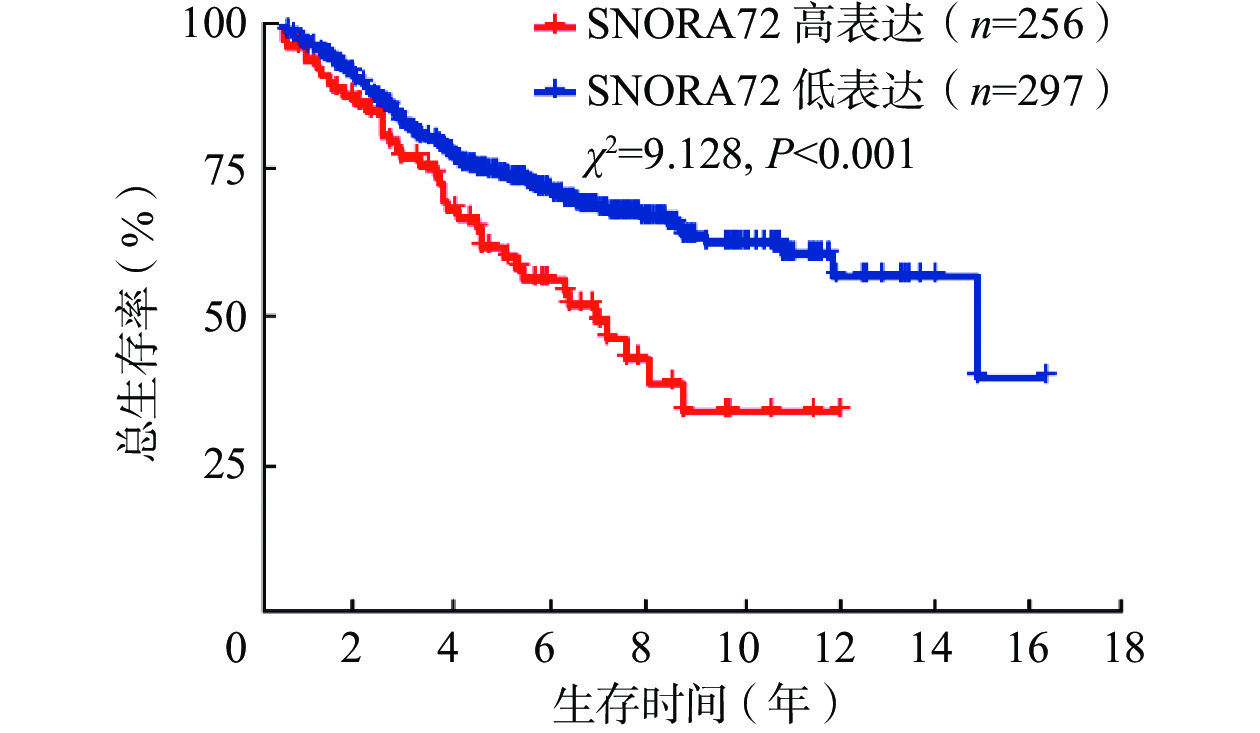
由表1可知,SNORA72表达与CRC患者年龄相关,且差异有统计学意义(P<0.05)。然后,进一步采用GSE39582数据集分析553例CRC患者SNORA72表达水平对患者预后的影响,结果显示,SNORA72高表达患者总生存率降低,且差异有统计学意义(χ2=9.128,P<0.001,图2)。这说明,SNORA72高水平表达可能是导致CRC患者临床预后不良的潜在危险因素。
临床特征 病例数 SNORA72表达 χ2值 P值 低表达 高表达 性别 0.143 0.706 女 29(46.7) 18(45.0) 11(50.0) 男 33(53.3) 22(55.0) 11(50.0) 年龄 5.350 0.021 ≤70岁 32(51.6) 25(62.5) 7(31.8) >70岁 30(48.4) 15(37.5) 15(68.2) TNM分期 2.741 0.433 Ⅰ /Ⅱ 44(71.0) 26(65.0) 18(81.8) Ⅲ/Ⅳ 18(29.0) 14(35.0) 4(18.2) 病理T分期 0.863 0.650 T1/T2 14(22.6) 8(20.0) 6(27.3) T3/T4 48(77.4) 32(80.0) 16(72.7) 淋巴结转移 1.851 0.396 阴性 45(72.6) 27(67.5) 18(81.8) 阳性 17(27.4) 13(32.5) 4(18.2) 远处转移 0.190 0.663 阴性 60(96.8) 39(97.5) 21(95.5) 阳性 2(3.2) 1(2.5) 1(4.5) 淋巴浸润 0.818 0.366 阴性 49(79.0) 33(82.5) 16(72.7) 阳性 13(21.0) 7(17.5) 6(27.3) 脉管浸润 0.845 0.358 阴性 54(87.1) 36(90.0) 18(81.8) 阳性 8(12.9) 4(10.0) 4(18.2) 结肠息肉史 1.097 0.295 阴性 52(83.9) 35(87.5) 17(77.3) 阳性 10(16.1) 5(12.5) 5(22.7) 注:TNM为肿瘤、淋巴结、转移 表 1 SNORA72表达与结直肠癌患者临床特征的相关性分析[例(%)]
Table 1. Correlation analysis of SNORA72 expression with clinical features of colorectal cancer patients (case (%))
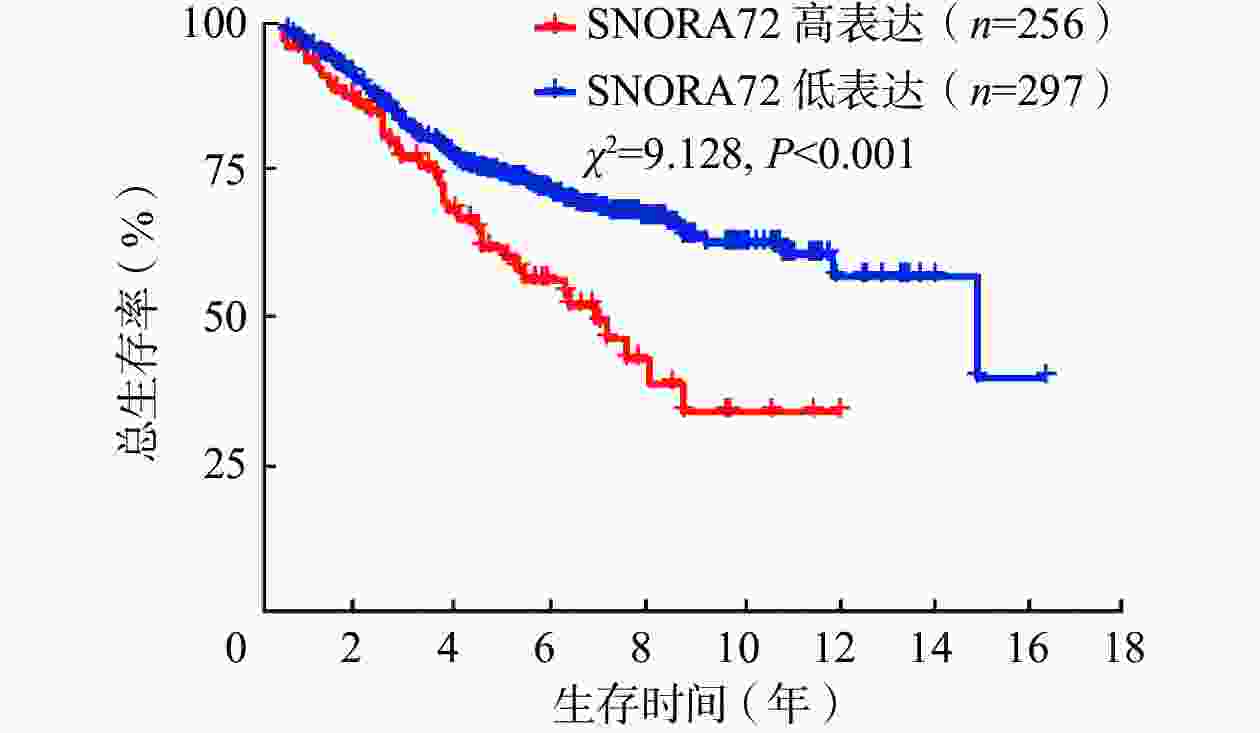
图 2 SNORA72表达影响结直肠癌患者预后的生存分析曲线(数据源于GSE39582数据集)
Figure 2. Survival analysis curves demonstrating the impact of SNORA72 expression on the prognosis of colorectal cancer patients
-
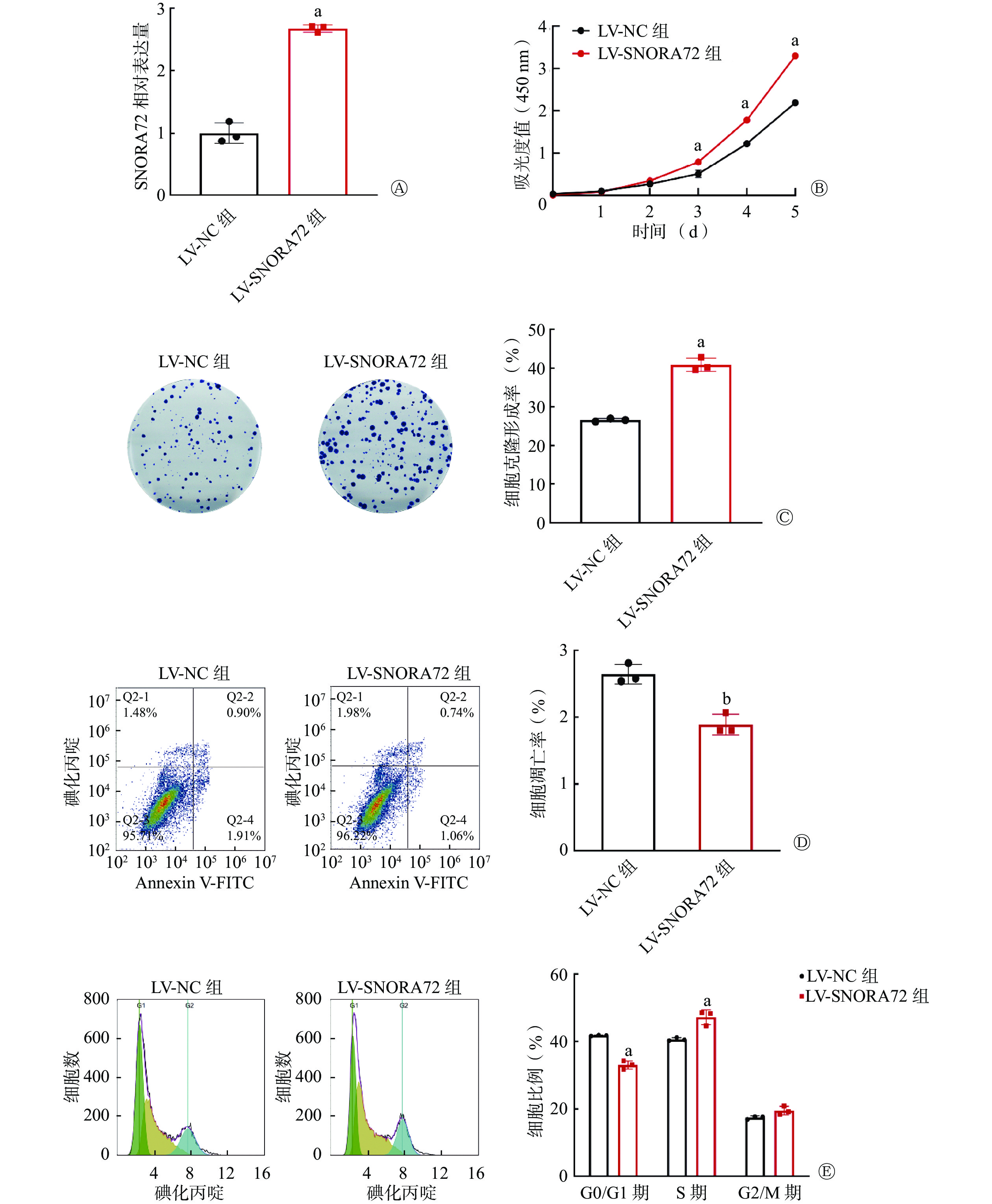
qRT-PCR 检测结果显示,与LV-NC组相比,LV-SNORA72组SNORA72的相对表达水平明显升高[(2.68±0.06)对(1.00±0.17)],且差异有统计学意义(t=16.570,P<0.001,图3A)。细胞增殖实验结果显示,在实验的第3、4、5天,LV-SNORA72组的吸光度值明显高于LV-NC组[(0.79±0.05)对(0.51±0.09)、(1.78±0.04)对(1.22±0.05)、(3.30±0.05)对(2.19±0.06)],且差异均有统计学意义(t=8.582、16.400、31.200,均P<0.001,图3B)。细胞克隆形成实验结果显示,LV-SNORA72组的细胞克隆形成率明显高于LV-NC组[(40.87±1.70)%对(26.60±0.40)%],且差异有统计学意义(t=14.140,P<0.001,图3C)。细胞凋亡检测结果显示,LV-SNORA72组的细胞凋亡率明显低于LV-NC组[(1.89±0.16)%对(2.64±0.15)%],且差异有统计学意义(t=6.115,P<0.01,图3D)。细胞周期分析结果显示,在G0/G1期,LV-SNORA72组的细胞比例明显低于LV-NC组[(33.00±1.23)%对(41.75±0.14)%],在S期,LV-SNORA72组的细胞比例明显高于LV-NC组[(47.16±2.18)%对(40.62±0.50)%],且差异均有统计学意义(t=9.099、6.806,均P<0.001,图3E)。以上实验结果表明,过表达SNORA72促进细胞增殖和克隆形成、抑制细胞凋亡,并通过改变细胞周期分布,减少了G1期细胞而增加了S期细胞的比例。

图 3 过表达SNORA72对结直肠癌细胞增殖、存活、凋亡和细胞周期分布的影响
Figure 3. Effect of overexpression of SNORA72 on proliferation, survival, apoptosis, and cell cycle distribution of colorectal cancer cells
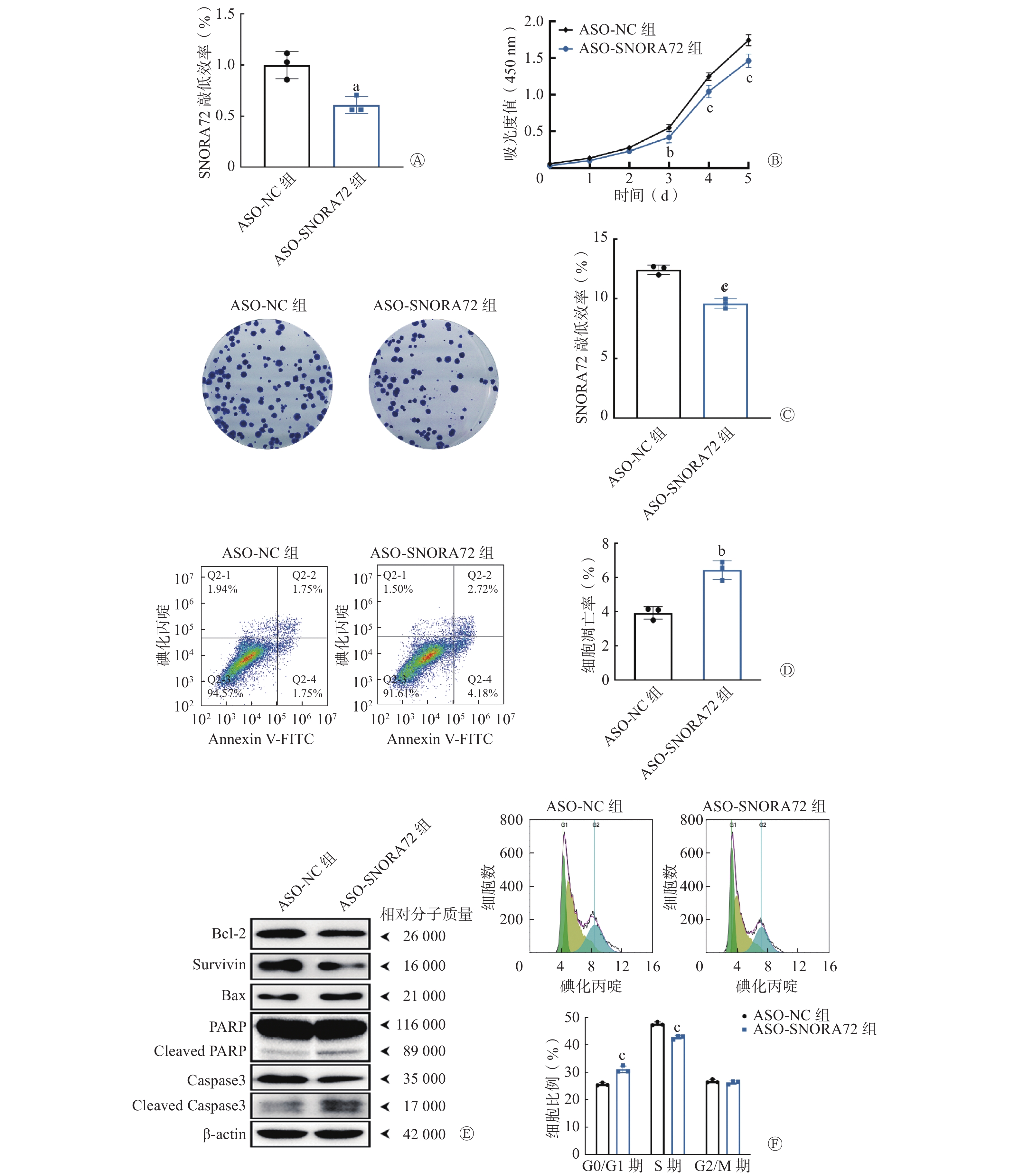
在此基础上,采用靶向SNORA72基因的ASO序列转染HT29细胞,进一步明确敲低SNORA72表达对细胞增殖、存活、凋亡和细胞周期分布的影响。qRT-PCR 检测结果显示,与ASO-NC组相比,ASO-SNORA72组SNORA72的相对表达水平明显降低[(0.61±0.08)对(1.00±0.13)],且差异有统计学意义(t=4.355,P<0.05,图4A)。细胞增殖实验结果显示,在实验的第3、4、5天,ASO-SNORA72组的吸光度值明显低于ASO-NC组[(0.42±0.07)对(0.55±0.05)、(1.04±0.08)对(1.25±0.05)、(1.46±0.09)对(1.74±0.08)],且差异有统计学意义(t=3.957、6.147、8.471,均P<0.01,图4B)。细胞克隆形成实验结果显示,ASO-SNORA72组的细胞克隆形成率明显低于ASO-NC组[(9.60±0.40)%对(12.43±0.38)%],且差异均有统计学意义(t=8.910,P<0.001,图4C)。细胞凋亡检测结果显示,ASO-SNORA72组的细胞凋亡率明显高于ASO-NC组[(6.44±0.54)%对(3.92±0.37)%],且差异有统计学意义(t=6.644,P<0.01,图4D)。Western blot实验结果显示,与ASO-NC组相比,ASO-SNORA72组可以促进凋亡蛋白PARP和Caspase3发生剪切活化,Bax蛋白表达增加,同时抑制抗凋亡蛋白Survivin和Bcl-2的表达(图4E)。另外,经细胞周期分析结果显示,在G0/G1期,ASO-SNORA72组的细胞比例明显高于ASO-NC组[(31.06±1.23)%对(25.59±0.55)%],在S期,ASO-SNORA72组的细胞比例明显低于ASO-NC组[(42.71±0.65)%对(47.71±0.67)%],且差异均有统计学意义(t=8.816、8.053,均P<0.001,图4F)。以上实验结果表明,敲低SNORA72表达可以抑制细胞增殖和克隆形成能力,并诱导细胞凋亡,同时导致细胞周期中G1期细胞比例增加而S期细胞比例减少。
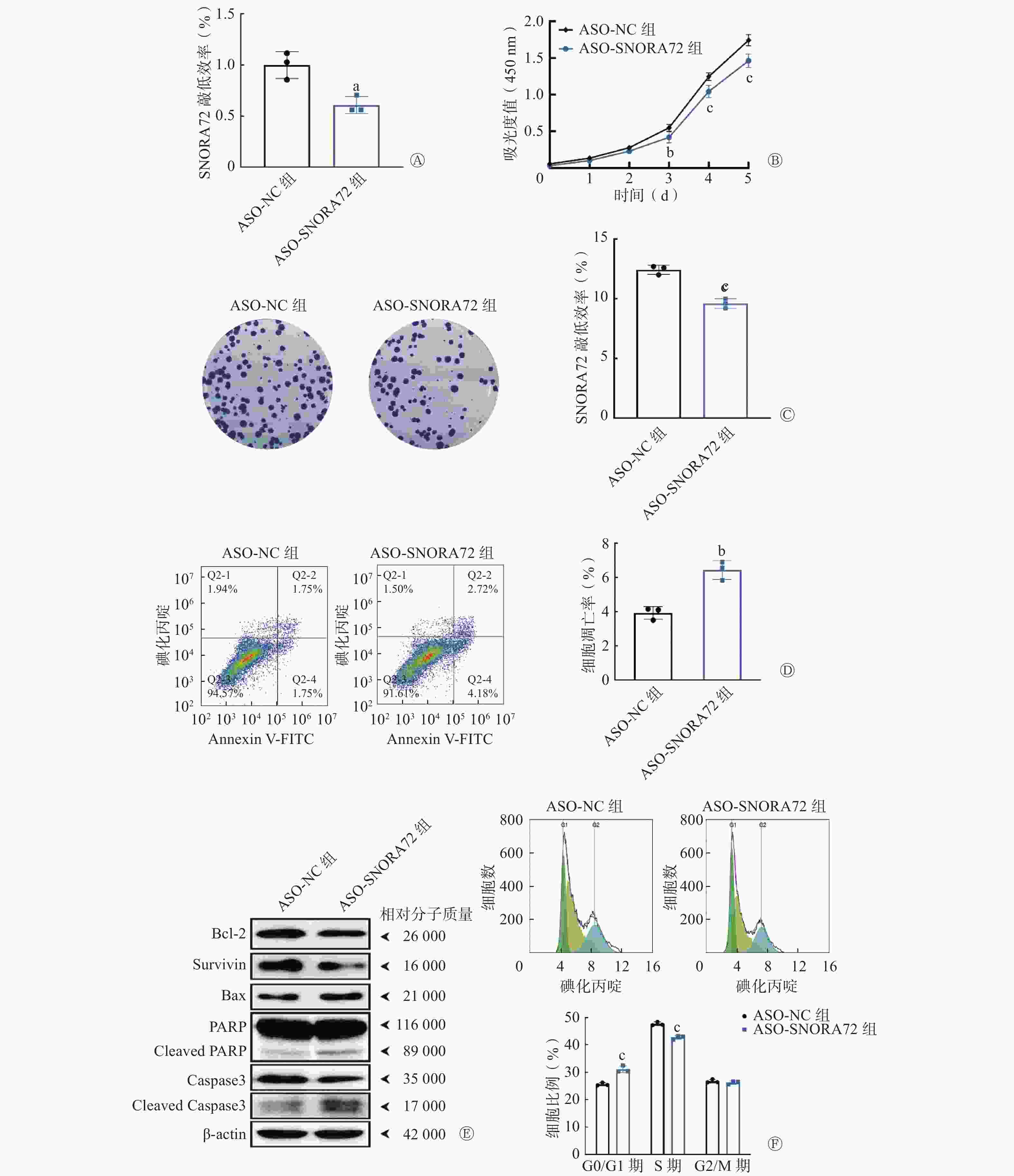
图 4 敲低SNORA72表达对结直肠癌细胞增殖、存活、凋亡和细胞周期分布的影响
Figure 4. Effect of knockdown of SNORA72 expression on proliferation, survival, apoptosis, and cycle distribution of colorectal cancer cells
-
放射敏感性分析的细胞克隆形成实验结果显示,在经1、2、4、6 Gy γ射线照射后,LV-SNORA72组的细胞存活分数均较LV-NC组增加[(0.89±0.05)对(0.81±0.03)、(0.64±0.10)对(0.47±0.01)、(0.16±0.04)对(0.09±0.01)、(0.04±0.01)对(0.02±0.01)],且差异均有统计学意义(t=4.063、8.802、4.045、2.937,均P<0.05,图5A)。由表2可知,过表达SNORA72可增强CRC细胞的放射抵抗性。
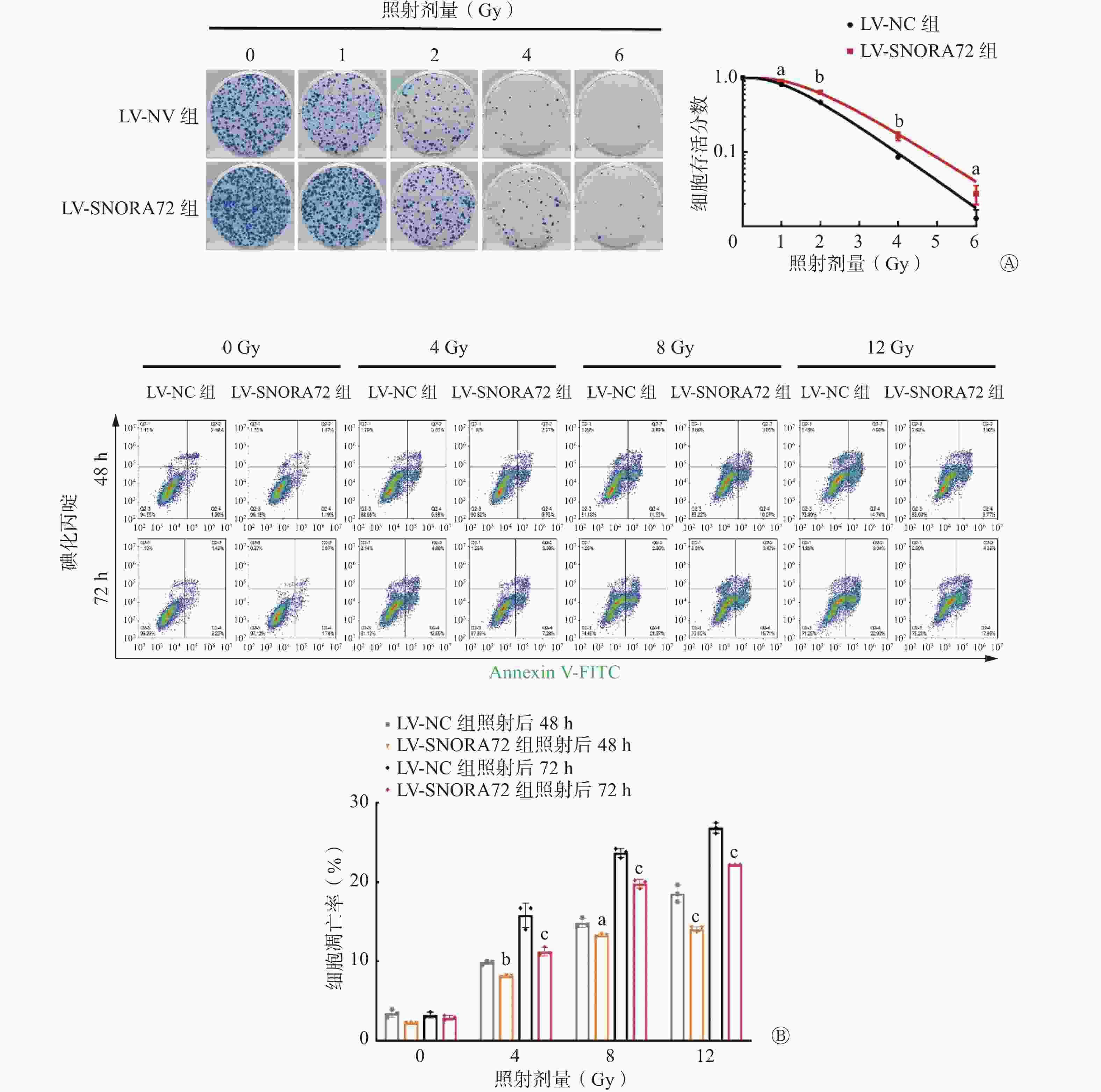
图 5 过表达SNORA72对结直肠癌细胞放射敏感性的影响
Figure 5. Effect of SNORA72 on radiosensitivity of colorectal cancer cells
组别 SF2 N D0(Gy) Dq(Gy) SER值 LV-NC组 0.47±0. 01 3.113 1.160 1.317 − LV-SNORA72组 0.64±0.10 3.911 1.312 1.789 0.736 注:−表示无此项数据。LV-NC组为过表达SNORA72阴性对照组;LV-SNORA72组为过表达SNORA72组;SF2为2 Gy照射后的细胞存活分数;N为外推值;D0为平均致死剂量;Dq为准阈剂量; SER为放射增敏比 表 2 基于集落形成分析SNORA72过表达后HT29细胞的单击-多靶模型参数值
Table 2. The parameters of a single-hit, multi-target model based on colony formation analysis of HT29 cells after SNORA72 overexpression
此外,放射诱导的细胞凋亡也是评价放射敏感性的重要指标。为了明确SNORA72是否通过调控细胞凋亡增加CRC细胞放射抵抗性,对LV-SNORA72组和LV-NC组细胞进行不同剂量 γ射线照射,结果显示,在4 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(8.14±0.12)%对(9.86±0.22)%、(11.26±0.52)%对(15.83±1.54)%],且差异均有统计学意义(t=3.470、9.208,均P<0.05);在8 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(13.29±0.17)%对(14.88±0.58)%、(19.82±0.56)%对(23.7±0.6)%],且差异均有统计学意义(t=3.201、7.819,均P<0.05);在12 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(14.06±0.32)%对(18.56±1.08)%、(22.19±0.02)%对(26.84±0.66)%],且差异均有统计学意义(t=9.054、9.369,均P<0.001,图5B)。以上实验结果表明,过表达SNORA72对放射诱导的细胞凋亡具有抑制作用。
-
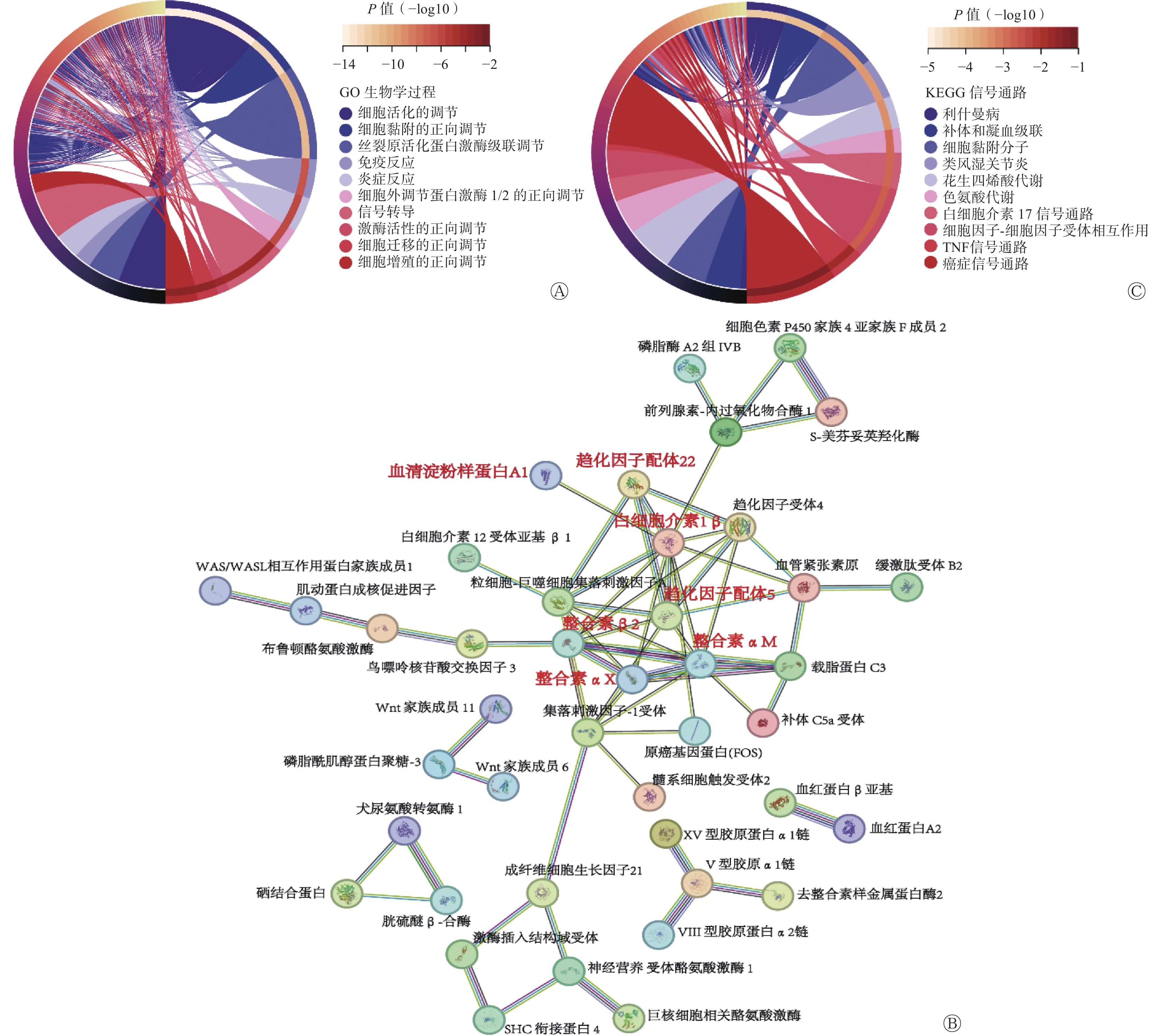
为了探究SNORA72调控CRC细胞生长和放射敏感性的分子机制,对LV-SNORA72和LV-NC感染的HT29细胞进行转录组RNA-seq测序分析。经数据归一化处理后,LV-NC组和LV-SNORA72组差异表达基因共筛选到815个差异性表达基因,包括638个上调基因和177个下调基因。基因本体论(GO)富集分析结果显示,SNORA72参与多种生物学过程,包括细胞活化和细胞黏附的调节、免疫和炎症反应、细胞迁移和增殖、细胞外调节蛋白激酶1/2(ERK1/2)和丝裂原活化蛋白激酶(MAPK)级联反应的正向调节等(图6A)。进一步对差异基因进行蛋白质-蛋白质间相互作用预测,我们发现了多种可能参与SNORA72功能调节的 “蛋白簇”,其中包括趋化因子配体5(CCL5)、白细胞介素1β(IL-1β)、趋化因子配体22(CCL22)、血清淀粉样蛋白A1(SAA1)等多种与细胞增殖、凋亡过程相关的炎症反应分子,整合素αM(ITGAM)、整合素β2(ITGB2)、整合素αX(ITGAX)等与细胞黏附、迁移过程相关的整合素家族分子(图6B)[12–17]。此外,对差异表达基因进行京都基因与基因组百科全书(KEGG)富集分析,结果显示过表达SNORA72细胞中,细胞黏附分子通路、癌症信号通路和炎症信号通路[肿瘤坏死因子(TNF)、白细胞介素17(IL-17)等]等多种与CRC疾病相关的信号通路被显著富集(图6C)。
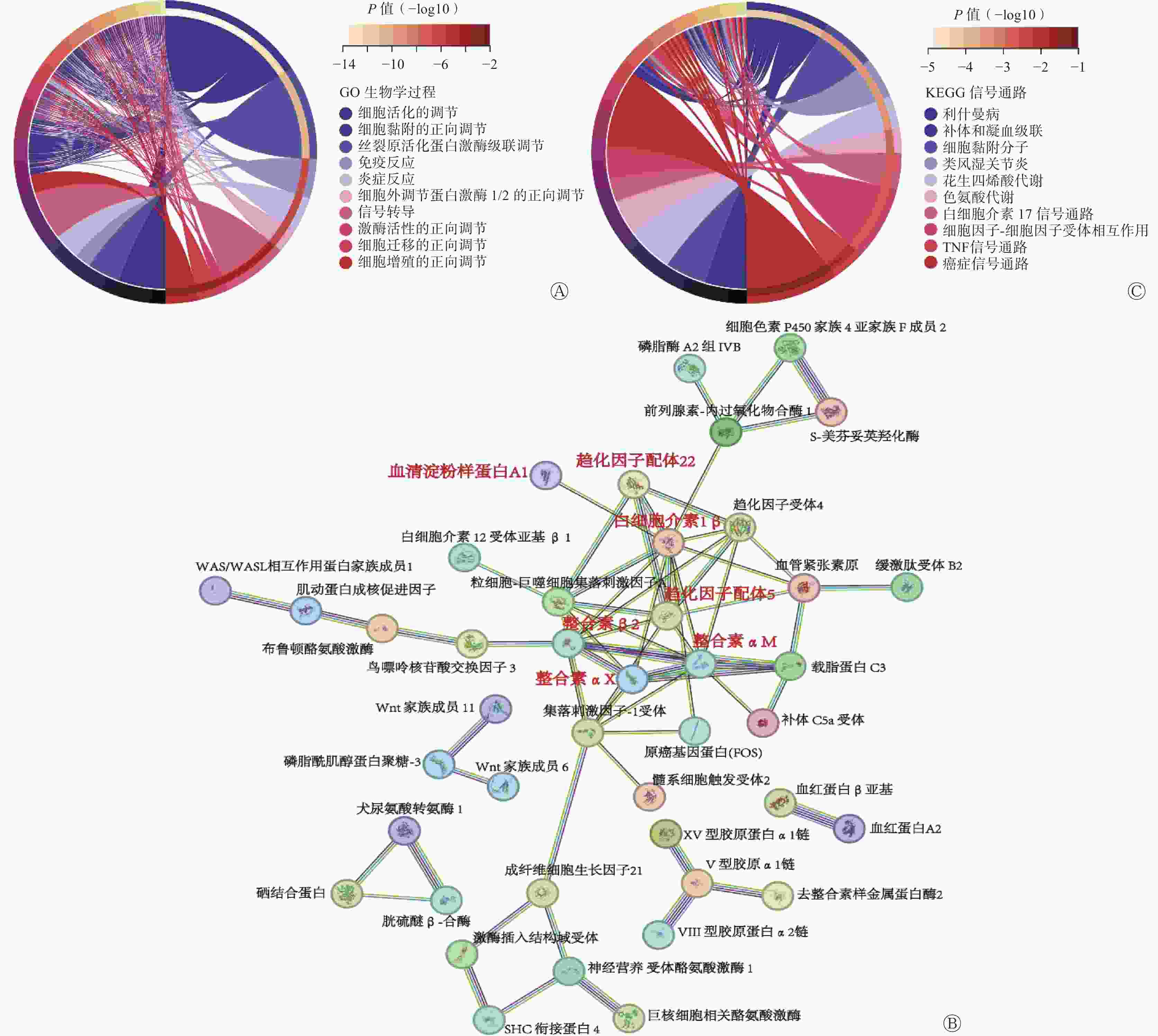
图 6 SNORA72靶向炎症反应及癌症相关信号通路
Figure 6. SNORA72 targets inflammatory response and cancer-related signaling pathways
-
近年来,snoRNA在癌症中的作用受到广泛关注,部分snoRNA被认为是潜在的恶性肿瘤标志物或治疗靶点[18]。但是,有关snoRNA对CRC疾病过程和放射敏感性作用及其临床应用潜能的研究仍然较少。目前已经发现一些snoRNA分子在CRC细胞中具有促癌作用,且表达水平与临床预后呈负相关[7–9]。特别值得关注的是,Liu等[19]发现snoRNA分子SNORD1C在CRC组织和患者血清中特异性高表达,其表达水平与血清中癌胚抗原水平具有显著相关性,被认为是CRC无创诊断的潜在生物标志物。在本研究中我们发现,snoRNA SNORA72在多种癌组织基因组中广泛扩增,其在CRC组织中的高水平表达与患者预后不良相关,提示其可作为CRC与癌旁组织分类诊断的候选标志物和预后评估新的潜在标志分子。Zhang等[10]报道,SNORA72在卵巢癌细胞中通过增强细胞的自我更新能力,促进癌细胞向干细胞特性的转化,从而发挥了促癌的功能。另外,在CRC细胞中,本研究结果证实SNORA72通过调控细胞周期、促进细胞增殖和细胞存活能力来发挥作用,这一发现进一步丰富了我们对这一分子的功能认识。但是,SNORA72生物学功能在不同组织细胞中是否具有普遍性目前仍然未知,需要进一步研究。
放疗是CRC临床治疗的重要手段,特别是近年来新辅助放疗在局部进展期直肠患者治疗中的应用,对于提高手术切除率、保肛率和延长生存期具有重要意义[20]。但是,由于个体间放射敏感性存在一定差异,部分患者对放疗不敏感,甚至完全抵抗,导致肿瘤复发和预后不良[21]。放射杀伤肿瘤细胞的基本原理是诱导细胞核DNA损伤,进而引发细胞凋亡或其他形式的细胞死亡。个别snoRNA分子,如SNORA73、scaRNA2可以与多聚二磷酸腺苷核糖聚合酶1(PARP-1)、DNA-依赖性蛋白激酶催化亚单位(DNA-PKcs)等重要DNA损伤修复分子发生直接作用,调节细胞DNA的修复能力[22-23],但是其是否参与细胞放射敏感性调控过程目前仍然未知。细胞克隆形成实验和细胞凋亡分析是评价细胞放射敏感性的经典实验方法[24-25],在本研究中,我们发现SNORA72能增加CRC细胞对γ射线的耐受能力,还发现snoRNA直接参与肿瘤细胞放射敏感性的调节。不足的是,本研究尚未探索SNORA72对肿瘤细胞放射敏感性的调节作用是否为SNORA72高表达的CRC患者预后不良的直接原因,及其是否对放射诱导的DNA损伤与修复过程产生影响。
据报道,snoRNA对肿瘤发病过程的调节作用往往与癌症相关信号通路活性异常相关,目前已经发现SNORD126、ACA11、SNORA71A等snoRNA能激活与肿瘤生长、侵袭、转移密切相关的磷脂酰肌醇3-激酶-AKT通路(PI3K-AKT)、丝裂原活化蛋白激酶/鸟苷酸激活蛋白激酶(MAPK/ERK)通路发挥促癌功能,而SNORD50A/B则能够抑制Ras-细胞外调节蛋白激酶1/2(Ras-ERK1/ERK2)通路活性发挥抑癌功能[26]。在结直肠细胞早期癌变过程中,癌基因[如Kirsten大鼠肉瘤病毒癌基因同源物(KRAS)、B-Raf蛋白(BRAF)]突变导致的MAPK/ERK通路活化是重要的遗传驱动因素[27]。在本研究中,我们通过转录组RNA-seq测序分析发现,SNORA72对MAPK/ERK信号级联反应具有正向调控作用,推测SNORA72可能靶向调控MAPK/ERK通路活性发挥促癌功能。另外,长期的慢性炎症反应是导致结直肠细胞癌变的重要原因[28]。我们还发现,过表达SNORA72 能显著富集肿瘤坏死因子、白细胞介素17介导的炎症信号通路及癌症信号通路,并且靶向调节多种炎症反应因子(趋化因子配体5、趋化因子配体22、白细胞介素1β、血清淀粉样蛋白A1等)。整合素则是一类能够调控肿瘤细胞侵袭、转移过程的黏附分子受体,在肿瘤恶性进展过程中起重要调控作用[29]。本研究中,我们发现整合素αM、整合素β2、整合素αX等整合素家族分子是SNORA72下游基因相互作用网络中的重要节点分子。但是,上述分子是否参与SNORA72 对CRC细胞的功能调控仍需后续进一步探索。
本研究结果显示,SNORA72在包括CRC在内的多种肿瘤组织中高表达,且其高表达与患者临床预后不良相关。该基因通过调节细胞G1/S期转换促进细胞增殖和克隆形成,抑制细胞凋亡,并能显著增强CRC细胞对放射损伤的抵抗性。本研究结果证实,SNORA72调节肿瘤细胞放射敏感性,在CRC中发挥促癌作用,具有成为CRC预后标志物的潜能。
利益冲突 所有作者声明无利益冲突
作者贡献声明 张文成负责实验的实施、数据的分析、论文的撰写;邓佳荣负责生物信息学的分析;刘鑫、张宏负责数据的收集与分析;王治东负责论文的修改;沈丽萍负责研究的设计、论文的修改与最终版本的修订
促癌基因SNORA72对结直肠癌细胞放射敏感性的作用研究
Oncogene SNORA72 enhances radioresistance in colorectal cancer cells
-
摘要:
目的 探索核仁小RNA(snoRNA)SNORA72基因在不同癌症特别是结直肠癌(CRC)中的表达模式及其对CRC细胞的生长及放射敏感性的影响。 方法 应用开放的癌症数据库分析SNORA72在不同癌症组织和CRC组织中的表达水平。构建过表达或敲低SNORA72的CRC细胞株HT29,将HT29细胞株分为过表达 SNORA72组(LV-SNORA72)及其阴性对照组(LV-NC)、敲低SNORA72表达组(ASO-SNORA72)及其阴性对照组(ASO-NC)。采用实时荧光定量聚合酶链反应(qRT-PCR)检测HT29细胞中SNORA72表达情况。分别检测在体外过表达或敲低SNORA72后,对细胞增殖、细胞克隆形成、细胞凋亡、细胞周期的影响。对LV-SNORA72组及LV-NC组的HT29细胞进行不同剂量60Co γ射线照射,检测各组细胞的存活分数(SF)和凋亡率。采用转录组学分析法探讨SNORA72影响HT29细胞生长可能的作用机制。两组间的比较采用独立样本t检验。 结果 癌症数据库分析发现SNORA72在包括CRC在内的多种癌症组织中高表达,且差异均有统计学意义(均P<0.05)。qRT-PCR 检测结果显示,与LV-NC组相比,LV-SNORA72组SNORA72的相对表达水平明显升高[(2.68±0.06)对(1.00±0.17)],且差异有统计学意义(t=16.570,P<0.001)。另外,与ASO-NC组相比,ASO-SNORA72组SNORA72的相对表达水平明显降低[(0.61±0.08)对(1.00±0.13)],且差异有统计学意义(t=4.355,P<0.05)。细胞增殖检测结果显示,在实验的第3、4、5天,LV-SNORA72组的吸光度值明显高于LV-NC组[(0.79±0.05)对(0.51±0.09)、(1.78±0.04)对(1.22±0.05)、(3.30±0.05)对(2.19±0.06)],且差异均有统计学意义(t=8.582、16.400、31.200,均P<0.001)。相反,ASO-SNORA72组的吸光度值明显低于ASO-NC组[(0.42±0.07)对(0.55±0.05)、(1.04±0.08)对(1.25±0.05)、(1.46±0.09)对(1.74±0.08)],且差异均有统计学意义(t=3.957、6.147、8.471,均P<0.01)。细胞克隆形成实验结果显示,LV-SNORA72组的克隆形成率明显高于LV-NC组[(40.87±1.70)%对(26.60±0.40)%],且差异有统计学意义(t=14.140,P<0.001)。相反,ASO-SNORA72组的克隆形成率明显低于ASO-NC组[(9.60±0.40)%对(12.43±0.38)%],且差异有统计学意义(t=8.910,P<0.001)。细胞凋亡检测结果显示,LV-SNORA72组的细胞凋亡率明显低于LV-NC组[(1.89±0.16)%对(2.64±0.15)%],且差异有统计学意义(t=6.115,P<0.01)。相反,ASO-SNORA72组的细胞凋亡率明显高于ASO-NC组[(6.44±0.54)%对(3.92±0.37)%],且差异有统计学意义(t=6.644,P<0.01)。Western blot结果显示,与ASO-NC组相比,ASO-SNORA72组可以促进凋亡蛋白PARP和Caspase3发生剪切活化,Bax蛋白表达水平升高,同时抑制抗凋亡蛋白Survivin和Bcl-2的表达。放射敏感性分析的细胞克隆形成实验结果显示,在经1、2、4、6 Gy γ射线照射后,LV-SNORA72组细胞的SF均较LV-NC组增加[(0.89±0.05)对(0.81±0.03)、(0.64±0.10)对(0.47±0.01)、(0.16±0.04)对(0.09±0.01)、(0.04±0.01)对(0.02±0.01)],且差异均有统计学意义(t=4.063、8.802、4.045、2.937,均P<0.05)。放射诱导的细胞凋亡结果显示,在4 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(8.14±0.12)%对(9.86±0.22)%、(11.26±0.52)%对(15.83±1.54)%],且差异均有统计学意义(t=3.470、9.208,均P<0.05);在8 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(13.29±0.17)%对(14.88±0.58)%、(19.82±0.56)%对(23.7±0.6)%],且差异均有统计学意义(t=3.201、7.819,均P<0.05);在12 Gy照射后48 h和72 h,与LV-NC组相比,LV-SNORA72组细胞凋亡率明显降低[(14.06±0.32)%对(18.56±1.08)%、(22.19±0.02)%对(26.84±0.66)%],且差异均有统计学意义(t=9.054、9.369,均P<0.001)。转录组学分析结果显示,SNORA72过表达影响细胞活化、细胞黏附、免疫和炎症反应,以及细胞迁移和增殖等生物学过程。 结论 SNORA72在CRC组织中特异性高表达且与患者不良预后相关,过表达SNORA72促进CRC细胞的生长和增殖,增加细胞放射抵抗性。 Abstract:Objective To explore the expression patterns of small nucleolar RNA (snoRNA) SNORA72 gene in various cancers, particularly in colorectal cancer (CRC), and its effect on the growth and radiosensitivity of CRC cells. Methods The expression of SNORA72 in different cancer and CRC tissues was analyzed using open cancer databases. The CRC cell line HT29, overexpressing or knocking down SNORA72, was constructed, dividing HT29 cells into the overexpressing SNORA72 group (LV-SNORA72) and its negative control group (LV-NC), as well as the SNORA72 knockdown group (ASO-SNORA72) and its negative control group (ASO-NC). The expression of SNORA72 in HT29 cells was detected by real-time fluorescence quantitative polymerase chain reaction (qRT-PCR). The effects of SNORA72 overexpression or knockdown on cell proliferation, colony formation, apoptosis, and cell cycle were evaluated. The HT29 cells from the LV-SNORA72 and LV-NC groups were irradiated with different doses of 60Co γ-rays, and the survival fraction (SF) and apoptosis rate of cells in each group were assessed. Transcriptomic analysis was employed to explore the potential mechanisms by which SNORA72 affects HT29 cell growth. An independent sample t-test was used for comparisons between two groups. Results Analysis of cancer databases revealed that SNORA72 is overexpressed in various cancer tissues, including CRC, the difference were statistically significant (all P<0.05). qRT-PCR results indicated that the relative expression of SNORA72 in the LV-SNORA72 group was significantly higher than that in the LV-NC group ((2.68±0.06) vs. (1.00±0.17)), and the difference was statistically significant (t=16.570, P<0.001). Conversely, the relative expression of SNORA72 in the ASO-SNORA72 group was significantly lower than that in the ASO-NC group ((0.61±0.08) vs. (1.00±0.13)), and the difference was statistically significant (t=4.355, P<0.05). Cell proliferation assay results showed that the absorbance values of the LV-SNORA72 group were significantly higher than those of the LV-NC group ((0.79±0.05) vs. (0.51±0.09), (1.78±0.04) vs. (1.22±0.05), and (3.30±0.05) vs. (2.19±0.06)) on the 3rd, 4th, and 5th day of the experiment, and the difference were statistically significant (t=8.582, 16.400, 31.200; all P<0.001). Conversely, the absorbance values of the ASO-SNORA72 group were significantly lower than those of the ASO-NC group ((0.42±0.07) vs. (0.55±0.05), (1.04±0.08) vs. (1.25±0.05), and (1.46±0.09) vs. (1.74±0.08)), and the difference were statistically significant (t=3.957, 6.147, 8.471; all P<0.01). Colony-formation assay results indicated that the colony formation rate of the LV-SNORA72 group was significantly higher than that of the LV-NC group((40.87±1.70)% vs. (26.60±0.40)%), and the difference was statistically significant (t=14.140, P<0.001). Conversely, the colony formation rate of the ASO-SNORA72 group was significantly lower than that of the ASO-NC group ((9.60±0.40)% vs. (12.43±0.38)%), and the difference was statistically significant (t=8.910, P<0.001). Apoptosis assay results showed that the apoptosis rate of the LV-SNORA72 group was significantly lower than that of the LV-NC group ((1.89±0.1)% vs. (2.64±0.15)%), and the difference was statistically significant (t=6.115, P<0.01). Conversely, the apoptosis rate of the ASO-SNORA72 group was significantly higher than that of the ASO-NC group((6.44±0.54)% vs. (3.92±0.37)%), and the difference was statistically significant (t=6.644, P<0.01). Western blot results demonstrated that compared with the ASO-NC group, the ASO-SNORA72 group promoted the cleavage activation of apoptosis proteins PARP and Caspase3, increased the expression of Bax protein, and inhibited the expression levels of anti-apoptotic proteins Survivin and Bcl-2. The results of radiosensitivity analysis through colony formation assay post-radiation showed that after exposure to 1, 2, 4, and 6 Gy of γ-rays, the SF of the LV-SNORA72 group increased compared with that of the LV-NC group ((0.89±0.05) vs. (0.81±0.03), (0.64±0.10) vs. (0.47±0.01), (0.16±0.04) vs. (0.09±0.01), and (0.04±0.01) vs. (0.02±0.01)), the difference were statistically significant (t=4.063, 8.802, 4.045, 2.937; all P<0.05). Radiation-induced apoptosis results showed that 48 h and 72 h after 4 Gy irradiation, the apoptosis rate in the LV-SNORA72 group was significantly lower than that in the LV-NC group ((8.14±0.12)% vs. (9.86±0.22)% and (11.26±0.52)% vs. (15.83±1.54%)), and the difference were statistically significant (t=3.470, 9.208; both P<0.05). After 8 Gy irradiation at 48 h and 72 h, the apoptosis rate in the LV-SNORA72 group was significantly lower than that in the LV-NC group ((13.29±0.17)% vs. (14.88±0.58)% and (19.82±0.56)% vs. (23.7±0.6)%), and the difference were statistically significant (t=3.201, 7.819; both P<0.05). After 12 Gy irradiation at 48 h and 72 h, the apoptosis rate in the LV-SNORA72 group was significantly lower than that in the LV-NC group ((14.06±0.32)% vs. (18.56±1.08)%) and (22.19±0.02)% vs. (26.84±0.66)%), the difference were statistically significant (t=9.054, 9.369; both P<0.001). Transcriptomic analysis results showed that overexpression of SNORA72 affects biological processes, such as cell activation, cell adhesion, and immune and inflammatory responses, cell migration, and cell proliferation. Conclusions SNORA72 is specifically overexpressed in CRC tissues and associated with poor prognosis in patients. It promotes CRC cell growth and proliferation and increases cellular radio-resistance. -
图 2 SNORA72表达影响结直肠癌患者预后的生存分析曲线(数据源于GSE39582数据集)
Figure 2. Survival analysis curves demonstrating the impact of SNORA72 expression on the prognosis of colorectal cancer patients
图 3 过表达SNORA72对结直肠癌细胞增殖、存活、凋亡和细胞周期分布的影响
Figure 3. Effect of overexpression of SNORA72 on proliferation, survival, apoptosis, and cell cycle distribution of colorectal cancer cells
图 4 敲低SNORA72表达对结直肠癌细胞增殖、存活、凋亡和细胞周期分布的影响
Figure 4. Effect of knockdown of SNORA72 expression on proliferation, survival, apoptosis, and cycle distribution of colorectal cancer cells
图 5 过表达SNORA72对结直肠癌细胞放射敏感性的影响
Figure 5. Effect of SNORA72 on radiosensitivity of colorectal cancer cells
图 6 SNORA72靶向炎症反应及癌症相关信号通路
Figure 6. SNORA72 targets inflammatory response and cancer-related signaling pathways
表 1 SNORA72表达与结直肠癌患者临床特征的相关性分析[例(%)]
Table 1. Correlation analysis of SNORA72 expression with clinical features of colorectal cancer patients (case (%))
临床特征 病例数 SNORA72表达 χ2值 P值 低表达 高表达 性别 0.143 0.706 女 29(46.7) 18(45.0) 11(50.0) 男 33(53.3) 22(55.0) 11(50.0) 年龄 5.350 0.021 ≤70岁 32(51.6) 25(62.5) 7(31.8) >70岁 30(48.4) 15(37.5) 15(68.2) TNM分期 2.741 0.433 Ⅰ /Ⅱ 44(71.0) 26(65.0) 18(81.8) Ⅲ/Ⅳ 18(29.0) 14(35.0) 4(18.2) 病理T分期 0.863 0.650 T1/T2 14(22.6) 8(20.0) 6(27.3) T3/T4 48(77.4) 32(80.0) 16(72.7) 淋巴结转移 1.851 0.396 阴性 45(72.6) 27(67.5) 18(81.8) 阳性 17(27.4) 13(32.5) 4(18.2) 远处转移 0.190 0.663 阴性 60(96.8) 39(97.5) 21(95.5) 阳性 2(3.2) 1(2.5) 1(4.5) 淋巴浸润 0.818 0.366 阴性 49(79.0) 33(82.5) 16(72.7) 阳性 13(21.0) 7(17.5) 6(27.3) 脉管浸润 0.845 0.358 阴性 54(87.1) 36(90.0) 18(81.8) 阳性 8(12.9) 4(10.0) 4(18.2) 结肠息肉史 1.097 0.295 阴性 52(83.9) 35(87.5) 17(77.3) 阳性 10(16.1) 5(12.5) 5(22.7) 注:TNM为肿瘤、淋巴结、转移  下载: 导出CSV
下载: 导出CSV
表 2 基于集落形成分析SNORA72过表达后HT29细胞的单击-多靶模型参数值
Table 2. The parameters of a single-hit, multi-target model based on colony formation analysis of HT29 cells after SNORA72 overexpression
组别 SF2 N D0(Gy) Dq(Gy) SER值 LV-NC组 0.47±0. 01 3.113 1.160 1.317 − LV-SNORA72组 0.64±0.10 3.911 1.312 1.789 0.736 注:−表示无此项数据。LV-NC组为过表达SNORA72阴性对照组;LV-SNORA72组为过表达SNORA72组;SF2为2 Gy照射后的细胞存活分数;N为外推值;D0为平均致死剂量;Dq为准阈剂量; SER为放射增敏比
下载: 导出CSV
-
[1] Zhang L, Cao F, Zhang GY, et al. Trends in and predictions of colorectal cancer incidence and mortality in China from 1990 to 2025[J/OL]. Front Oncol, 2019, 9: 98[2023-10-08]. https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2019.00098/full. DOI: 10.3389/fonc.2019.00098. [2] Dekker E, Tanis PJ, Vleugels JLA, et al. Colorectal cancer[J]. Lancet, 2019, 394(10207): 1467−1480. DOI: 10.1016/S0140-6736(19)32319-0. [3] Siegel RL, Miller KD, Goding Sauer A, et al. Colorectal cancer statistics, 2020[J]. CA Cancer J Clin, 2020, 70(3): 145−164. DOI: 10.3322/caac.21601. [4] Diagnosis And Treatment Guidelines For Colorectal Cancer Working Group CSOCOC. Chinese Society of Clinical Oncology (CSCO) diagnosis and treatment guidelines for colorectal cancer 2018 (English version)[J/OL]. Chin J Cancer Res, 2019, 31(1): 117−134[2023-10-08]. DOI: 10.21147/j.issn.1000-9604.2019.01.07. [5] Patra Bhattacharya D, Canzler S, Kehr S, et al. Phylogenetic distribution of plant snoRNA families[J]. BMC Genom, 2016, 17(1): 969. DOI: 10.1186/s12864-016-3301-2. [6] Williams GT, Farzaneh F. Are snoRNAs and snoRNA host genes new players in cancer?[J]. Nat Rev Cancer, 2012, 12(2): 84−88. DOI: 10.1038/nrc3195. [7] Yoshida K, Toden S, Weng WH, et al. SNORA21−an oncogenic small nucleolar RNA, with a prognostic biomarker potential in human colorectal cancer[J/OL]. EBioMedicine, 2017, 22: 68−77[2023-10-08]. https://www.thelancet.com/journals/ebiom/article/PIIS2352-3964(17)30282-7/fulltext. DOI: 10.1016/j.ebiom.2017.07.009. [8] Okugawa Y, Toiyama Y, Toden S, et al. Clinical significance of SNORA42 as an oncogene and a prognostic biomarker in colorectal cancer[J]. Gut, 2017, 66(1): 107−117. DOI: 10.1136/gutjnl-2015-309359. [9] Zhang ZX, Tao YX, Hua QL, et al. SNORA71A promotes colorectal cancer cell proliferation, migration, and invasion[J/OL]. Biomed Res Int, 2020, 2020: 8284576[2023-10-08]. https://www.hindawi.com/journals/bmri/2020/8284576. DOI: 10.1155/2020/8284576. [10] Zhang LW, Ma R, Gao MC, et al. SNORA72 activates the notch1/c-Myc pathway to promote stemness transformation of ovarian cancer cells[J/OL]. Front Cell Dev Biol, 2020, 8: 583087[2023-10-08]. https://www.frontiersin.org/articles/10.3389/fcell.2020.583087/full. DOI: 10.3389/fcell.2020.583087. [11] Myllykangas S, Böhling T, Knuutila S. Specificity, selection and significance of gene amplifications in cancer[J]. Semin Cancer Biol, 2007, 17(1): 42−55. DOI: 10.1016/j.semcancer.2006.10.005. [12] 吴颖, 高蔚, 张佑杨, 等. CCL5对人肺腺癌细胞增殖、凋亡的影响及机制研究[J]. 淮海医药, 2022, 40(6): 565−569. DOI: 10.14126/j.cnki.1008-7044.2022.06.004.
Wu Y, Gao W, Zhang YY, et al. Effects of CCL5 on proliferation, migration and apoptosis of lung cancer A549 cells[J]. J Huaihai Med, 2022, 40(6): 565−569. DOI: 10.14126/j.cnki.1008-7044.2022.06.004.[13] CNTF protects MIN6 cells against apoptosis induced by Alloxan and IL-1β through downregulation of the AMPK pathway[J]. Cell Signal, 2011, 23(10): 1669-1676. DOI:10.1016/j.cellsig.2011.06.001. [14] Zhou M, Wang X, Shi YQ, et al. Deficiency of ITGAM attenuates experimental abdominal aortic aneurysm in mice[J/OL]. J Am Heart Assoc, 2021, 10(7): e019900[2023-10-08]. https://www.ahajournals.org/doi/10.1161/JAHA.120.019900. DOI: 10.1161/JAHA.120.019900. [15] Xu ZJ, Wu YF, Fu GH, et al. SAA1 has potential as a prognostic biomarker correlated with cell proliferation, migration, and an indicator for immune infiltration of tumor microenvironment in clear cell renal cell carcinoma[J/OL]. Int J Mol Sci, 2023, 24(8): 7505[2023-10-08]. https://www.mdpi.com/1422-0067/24/8/7505. DOI: 10.3390/ijms24087505. [16] Guo W, Cai YL, Liu XM, et al. Single-exosome profiling identifies ITGB3+ and ITGAM+ exosome subpopulations as promising early diagnostic biomarkers and therapeutic targets for colorectal cancer[J/OL]. Research (Wash D C), 2023, 6: 0041[2023-10-08]. https://spj.science.org/doi/10.34133/research.0041. DOI: 10.34133/research.0041. [17] Chen J, Zhao D, Zhang LY, et al. Tumor-associated macrophage (TAM)-derived CCL22 induces FAK addiction in esophageal squamous cell carcinoma (ESCC)[J]. Cell Mol Immunol, 2022, 19(9): 1054−1066. DOI: 10.1038/s41423-022-00903-z. [18] Stepanov GA, Filippova JA, Komissarov AB, et al. Regulatory role of small nucleolar RNAs in human diseases[J/OL]. Biomed Res Int, 2015, 2015: 206849[2023-10-08]. https://www.hindawi.com/journals/bmri/2015/206849. DOI: 10.1155/2015/206849. [19] Liu YH, Zhao CW, Sun J, et al. Overexpression of small nucleolar RNA SNORD1C is associated with unfavorable outcome in colorectal cancer[J]. Bioengineered, 2021, 12(1): 8943−8952. DOI: 10.1080/21655979.2021.1990194. [20] Thariat J, Hannoun-Levi JM, Sun Myint A, et al. Past, present, and future of radiotherapy for the benefit of patients[J]. Nat Rev Clin Oncol, 2013, 10(1): 52−60. DOI: 10.1038/nrclinonc.2012.203. [21] Benson AB, Venook AP, Al-Hawary MM, et al. NCCN guidelines insights: colon cancer, version 2.2018[J]. J Natl Compr Canc Netw, 2018, 16(4): 359−369. DOI: 10.6004/jnccn.2018.0021. [22] Han C, Sun LY, Luo XQ, et al. Chromatin-associated orphan snoRNA regulates DNA damage-mediated differentiation via a non-canonical complex[J/OL]. Cell Rep, 2022, 38(13): 110421[2023-10-08]. https://www.sciencedirect.com/science/article/pii/S2211124722001450. DOI: 10.1016/j.celrep.2022.110421. [23] Bergstrand S, O'Brien EM, Coucoravas C, et al. Small Cajal body-associated RNA 2 (scaRNA2) regulates DNA repair pathway choice by inhibiting DNA-PK[J/OL]. Nat Commun, 2022, 13(1): 1015[2023-10-08]. https://www.nature.com/articles/s41467-022-28646-5. DOI: 10.1038/s41467-022-28646-5. [24] Shen LF, Zhou SH, Yu Q. Predicting response to radiotherapy in tumors with PET/CT: when and how?[J]. Transl Cancer Res, 2020, 9(4): 2972−2981. DOI: 10.21037/tcr.2020.03.16. [25] Zhivotovsky B, Joseph B, Orrenius S. Tumor radiosensitivity and apoptosis[J]. Exp Cell Res, 1999, 248(1): 10−17. DOI: 10.1006/excr.1999.4452. [26] Siprashvili Z, Webster DE, Johnston D, et al. The noncoding RNAs SNORD50A and SNORD50B bind K-Ras and are recurrently deleted in human cancer[J]. Nat Genet, 2016, 48(1): 53−58. DOI: 10.1038/ng.3452. [27] Fang JY, Richardson BC. The MAPK signalling pathways and colorectal cancer[J]. Lancet Oncol, 2005, 6(5): 322−327. DOI: 10.1016/S1470-2045(05)70168-6. [28] Schmitt M, Greten FR. The inflammatory pathogenesis of colorectal cancer[J]. Nat Rev Immunol, 2021, 21(10): 653−667. DOI: 10.1038/s41577-021-00534-x. [29] Hamidi H, Ivaska J. Every step of the way: integrins in cancer progression and metastasis[J]. Nat Rev Cancer, 2018, 18(9): 533−548. DOI: 10.1038/s41568-018-0038-z. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4103
- HTML全文浏览量: 2820
- PDF下载量: 2
