


 下载:
下载:

-
氚是氢的同位素之一,随着核技术的应用与发展,核试验、核反应堆、核燃料后处理、核事故的发生等会导致大量的氚排放到环境中[1-3]。氚的理化性质决定了其在环境中大多以氚水的形式存在,且可以经皮肤、伤口、呼吸道、消化道等多种途径进入生物体内造成内照射损伤[4]。高剂量氚的相对生物效能(relative biological effectiveness, RBE)值为1[5],但近年来的一些研究结果表明,低剂量氚(<100 mGy)的RBE值可能>1,且RBE值在一定范围内随氚水剂量的降低而升高[6-7]。低剂量氚水所致的生物效应不易观察,需要通过构建合适的动物模型才可以检测[8]。
斑马鱼是一种模式生物,其具有体型小、养成周期短、产卵量大、胚胎透明、成本低等优点[9],是国际标准化组织(ISO)认可的5种鱼类实验动物之一,被经济合作与发展组织(OECD)推荐用于各种类型的生态毒理学试验[10-11]。
本研究拟通过斑马鱼构建氚水长期暴露动物模型,并在此基础上检测斑马鱼子代发生的改变[12],初步探讨斑马鱼长期氚水暴露可能导致的子代生物效应。
-
实验所用成年AB品系野生型斑马鱼购自国家斑马鱼资源中心。本实验经苏州大学动物保护和使用委员会批准,符合《实验动物护理和使用指南》的要求。
-
氚水购自美国PerkinElmer公司;培养基(5 mmol/L NaCl,0.17 mmol/L KCl,0.33 mmol/L CaCl2,0.03 mmol/L MgSO4,0.01%亚甲基蓝和1 L去离子水)为本实验室自行配制;PBS购自美国AXYFEN公司;总超氧化物歧化酶(total superoxide dismutase,T-SOD)测定试剂盒、丙二醛(malondialdehyde,MDA)测定试剂盒、活性氧(reactive oxygen species,ROS)测定试剂盒均购自南京建成生物工程研究所;三卡因(Tricaine)购自苏州格瑞特医药技术有限公司;高氯酸购自上海阿拉丁生化科技股份有限公司;过氧化氢购自上海凌峰化学试剂有限公司。5 ml可立冻存管购自苏州科技有限公司;斑马鱼养殖系统购自青岛金水海洋生物设备有限公司;低本底液闪计数器(Tri-Carb 2910TR 型)购自美国PerkinElmer公司;MoticSMZ-168型体式显微镜购自厦门麦克奥迪实业集团有限公司;十二孔细菌培养板购自上海赛默尔世飞科技(中国)有限公司;MF52-N型荧光显微镜购自广州明美光电技术有限公司;KZ-Ⅲ型研磨仪购自武汉赛维尔生物科技有限公司;DK-S22型电热恒温水浴锅购自上海精宏实验设备有限公司。
-
将正常斑马鱼所产胚胎分别暴露于0、1×102、1×105 Bq/L的氚水中3个月以上作为亲代(记作F0代),待其性成熟后进行繁殖得到子代(记作F1代)并继续饲养在对应浓度的氚水中。按照斑马鱼的发育阶段,分别选取合适的指标对胚胎期、幼苗期、幼鱼期、成鱼期斑马鱼进行相关检测。
-
采用简单随机方法分别从0、1×102、1×105 Bq/L的3组氚水暴露F0代斑马鱼成鱼所产胚胎中选取50枚胚胎,继续暴露在与F0代对应的0、1×102、1×105 Bq/L氚水中,第7天时统计每组50枚胚胎的孵化数并计算孵化率(孵化率=孵化的斑马鱼数量/50×100%),期间及时去除死卵。上述实验至少重复3次。
-
在F1代受精后24、36 h,采用简单随机方法分别从1.3.2中的3组F1代斑马鱼胚胎中各选取10枚胚胎,使用体式显微镜对斑马鱼胚胎1 min内的自主运动次数计数;受精后48、60 h,采用简单随机方法分别从上述3组胚胎中各选取10枚胚胎,使用体式显微镜对斑马鱼胚胎20 s内的心脏跳动次数计数;受精后72、84 h,采用简单随机方法分别从上述3组斑马鱼胚胎中各选取10条斑马鱼幼苗,在体式显微镜下拍照,应用麦克奥迪图像软件(Motic Images Plus,厦门麦克奥迪实业集团有限公司)计算3组斑马鱼幼苗的体长。上述实验均至少重复3次。
-
将F0代3组斑马鱼所产胚胎继续暴露于与F0代对应的0、1×102、1×105 Bq/L 3组不同浓度的氚水中,在斑马鱼胚胎受精4~5 h时加入0.003% 苯基硫脲(PTU)溶液以抑制斑马鱼体内黑色素的生成。在斑马鱼幼苗鱼龄达4 d时,采用简单随机方法从0、1×102、1×105 Bq/L的3个浓度氚水暴露组中各选取10条斑马鱼鱼苗进行实验。每组斑马鱼鱼苗分别加入12孔板中,用PBS清洗3次,随后加入1 ml含10 μmol/L DCFH-DA(2, 7-二氯荧光素二乙酸酯)的PBS,于28.5℃、避光环境中孵育1 h。孵育结束后,用PBS清洗3次以去除多余的探针,在荧光显微镜下拍照记录3组斑马鱼体内的ROS荧光强度。采用简单随机方法从3组中分别选取4条斑马鱼,采用Image J软件(美国NIH公司)对其全身的荧光强度进行定量分析。
-
采用简单随机方法选取F1代0、1×102、1×105 Bq/L 3个浓度氚水暴露组斑马鱼幼鱼中鱼龄达45、60 d的各3条,准确称量体重后按照体重(g)∶生理盐水(ml)=1∶9的比例在研磨仪上研磨制成10%的组织匀浆,再使用生理盐水稀释为5%的组织匀浆。吸取5%的组织匀浆30 μl,使用生理盐水稀释为1%的组织匀浆检测T-SOD的含量;吸取5%的组织匀浆100 μl,使用二喹啉甲酸(BCA)法对蛋白浓度进行定量分析;吸取5%的组织匀浆100 μl检测MDA的含量。实验步骤均严格按照对应试剂盒的说明书进行。
-
采用简单随机方法分别选取0、1×102、1×105 Bq/L 3个浓度氚水暴露组中4月龄以上性成熟的F1代斑马鱼雌、雄鱼各1条,在交配前1天晚上放入配鱼缸,中间用隔板隔开。第二天上午9点,自动光照系统(本实验室自制)开启后将隔板拿开,雌鱼开始产卵2 h后收集斑马鱼胚胎并计数。上述实验至少重复3次。
-
采用简单随机方法分别选取0、1×102、1×105 Bq/L 3个浓度氚水暴露组中鱼龄为45、60 d的斑马鱼幼鱼各3条,使用三卡因将其麻醉后用纯净水冲洗3次,擦干幼鱼体表水分后称量体重,按照体重(g)∶生理盐水(ml)=1∶9的比例在研磨仪上研磨制成10%的组织匀浆。吸取10%的组织匀浆500 μl于5 ml可立冻存管中,加入500 μl消解液(HClO4∶H2O2=2∶3),在70℃电热恒温水浴锅中消解至少1 h,待消解完全后吸取全部液体于20 ml液闪瓶中,加入7 ml纯净水混合均匀。对照组为8 ml纯净水。在避光条件下,向液闪瓶中加入12 ml闪烁液并充分振荡,加盖密封暗化12 h以上至液体澄清后,使用低本底液闪计数器对斑马鱼体内的总氚含量进行检测。检测过程全程避光,检测时间10 min,检测设置内循环1次,外循环3次。
-
采用Graphpad Prism 9软件对数据进行统计学分析。符合正态分布的计量资料以
$ \bar{x}\pm s $ 表示,两组间的比较采用t检验(方差齐)。P<0.05为差异有统计学意义。 -
由图1可知,0、1×102、1×105 Bq/L氚水暴露组斑马鱼的孵化率分别为(90.66±0.05)%、(85.63±0.10)%、(78.06±0.15)%。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼孵化率的差异均无统计学意义(t=0.785、1.370,P=0.462、0.220)。

图 1 不同浓度氚水长期暴露F1代斑马鱼孵化率的比较
Figure 1. Comparison of hatching rate of F1 generation zebrafish after long-term exposure to different concentrations of tritiated water
-
由图2可知,0、1×102、1×105 Bq/L氚水暴露组斑马鱼受精后24 h的自主运动次数分别为(12.93±2.70)、(11.30±0.78)、(10.50±0.80) 次/min。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后24 h自主运动次数的差异均无统计学意义(t=1.008、1.499,P=0.370、0.208)。3组斑马鱼受精后36 h的自主运动次数分别为(3.63±1.43)、(4.50±1.15)、(5.40±3.55) 次/min。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后36 h自主运动次数的差异均无统计学意义(t=0.817、0.799,P=0.460、0.469)。

图 2 不同浓度氚水长期暴露F1代斑马鱼受精后不同时间自主运动次数、心率、体长的比较
Figure 2. Comparison of autonomous movement, heart rate and body length of F1 generation zebrafish at different times after fertilization after long-term exposure to different concentrations of tritiated water
3组斑马鱼受精后48 h的心率分别为(59.43±6.93)、(65.00±3.30)、(61.23±4.55) 次/20 s。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后48 h心率的差异均无统计学意义(t=1.256、0.376,P=0.278、P=0.726)。3组斑马鱼受精后60 h的心率分别为(69.87±2.71)、(66.17±6.97)、(69.77±9.08) 次/20 s。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后60 h心率的差异均无统计学意义(t=0.857、0.018,P=0.440、0.986)。
3组斑马鱼受精后72 h的体长分别为(3.20±0.22)、(3.32±0.08)、(3.29±0.06) mm。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后72 h体长的差异均无统计学意义(t=0.614、0.178,P=0.525、0.868)。3组斑马鱼受精后84 h的体长分别为(3.42±0.07)、(3.46±0.11)、(3.40±0.04) mm。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后84 h体长的差异均无统计学意义(t=0.527、0.496,P=0.626、0.646)。
-
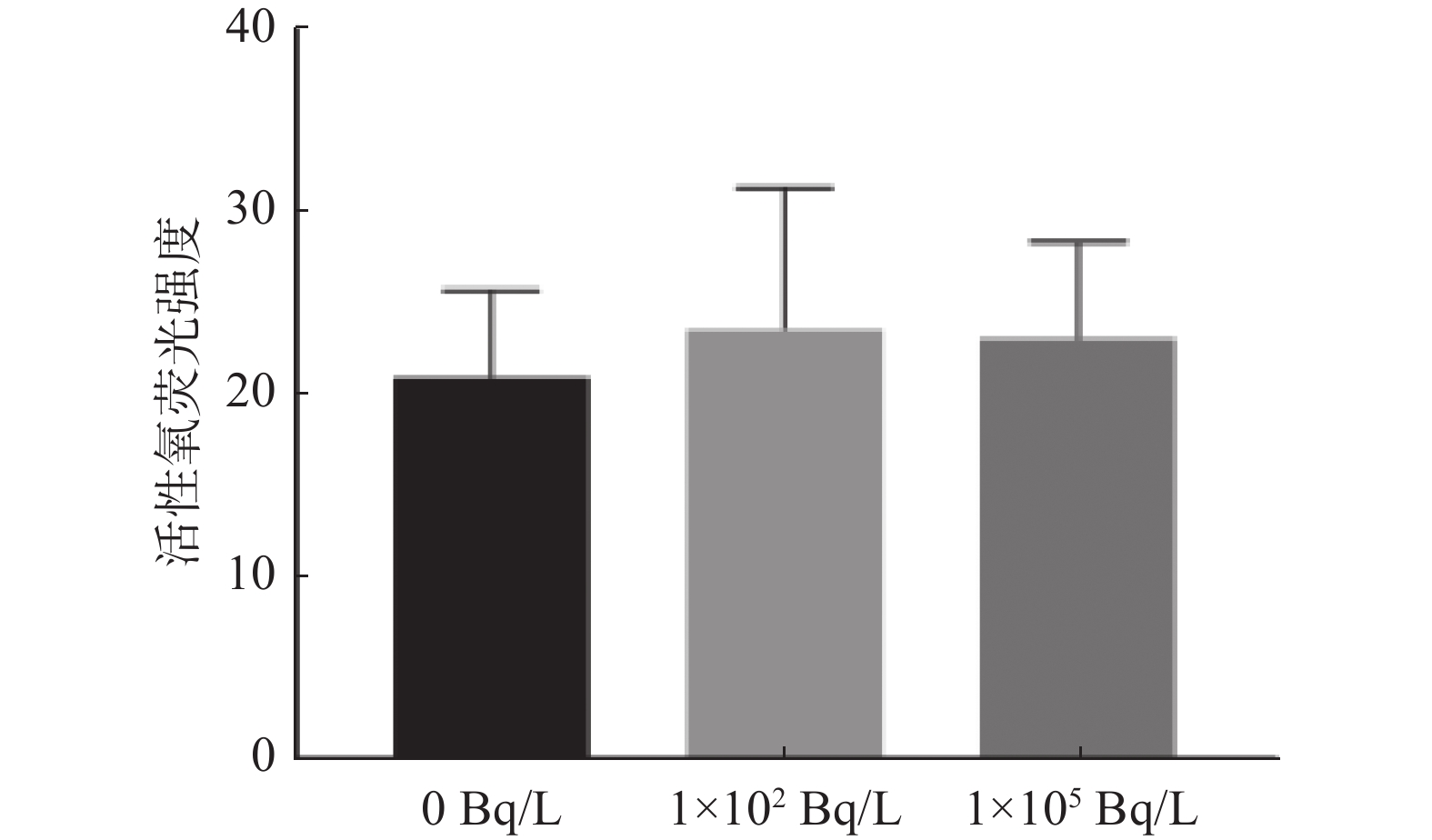
由图3可知,0、1×102、1×105 Bq/L氚水暴露组斑马鱼的ROS荧光强度分别为(21.07±4.74)、(23.71±7.73)、(23.19±5.32)。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼幼苗ROS荧光强度的差异均无统计学意义(t=0.582、0.593,P=0.582、0.575)。

图 3 不同浓度氚水长期暴露F1代斑马鱼幼苗活性氧荧光强度的比较
Figure 3. Comparison of reactive oxygen species fluorescence intensity of F1 generation zebrafish seedlings after long-term exposure to different concentrations of tritiated water
-
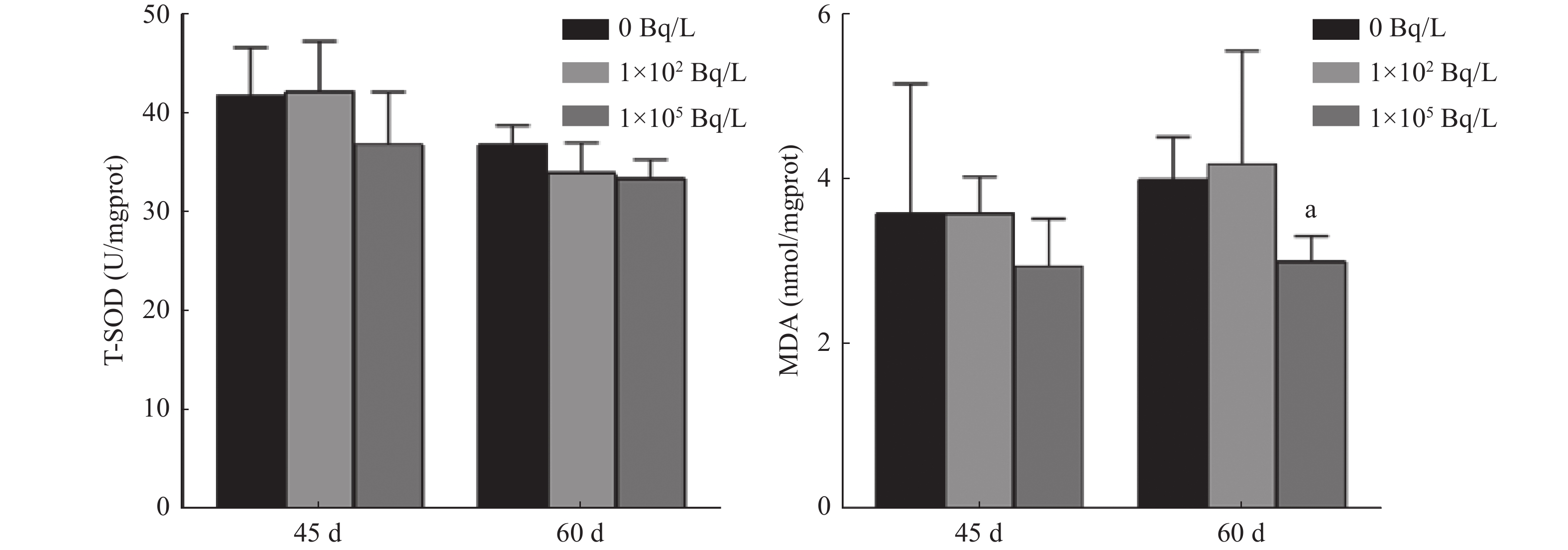
由图4可知,0、1×102、1×105 Bq/L氚水暴露组斑马鱼45 d的T-SOD含量分别为(41.84±4.91)、(42.30±5.04)、(36.97±5.26) U/mgprot。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼45 d T-SOD含量的差异均无统计学意义(t=0.112、1.171,P=0.916、0.307)。3组斑马鱼60 d的T-SOD含量分别为(36.93±1.91)、(34.07±3.02)、(33.54±1.87) U/mgprot。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼60 d T-SOD含量的差异均无统计学意义(t=1.397、2.195,P=0.240、0.093)。

图 4 不同浓度氚水长期暴露F1代不同鱼龄斑马鱼幼鱼体内T-SOD、MDA含量的比较
Figure 4. Comparison of total superoxide dismutase and malondialdehyde content in F1 generation zebrafish of different age after long-term exposure to different concentrations of tritiated water
3组斑马鱼45 d的MDA含量分别为(3.60±1.56)、(3.59±0.44)、(2.95±0.58) nmol/mgprot。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼45 d MDA含量的差异均无统计学意义(t=0.007、0.677,P=0.995、0.536)。3组斑马鱼60 d的MDA含量分别为(4.00±0.52)、(4.19±1.37)、(3.01±0.32) nmol/mgprot。与0 Bq/L氚水暴露组相比,1×102 Bq/L氚水暴露组F1代斑马鱼60 d MDA含量的差异无统计学意义(t=0.229,P=0.830),1×105 Bq/L氚水暴露组F1代斑马鱼MDA含量的差异有统计学意义(t=2.831,P=0.047)。
-
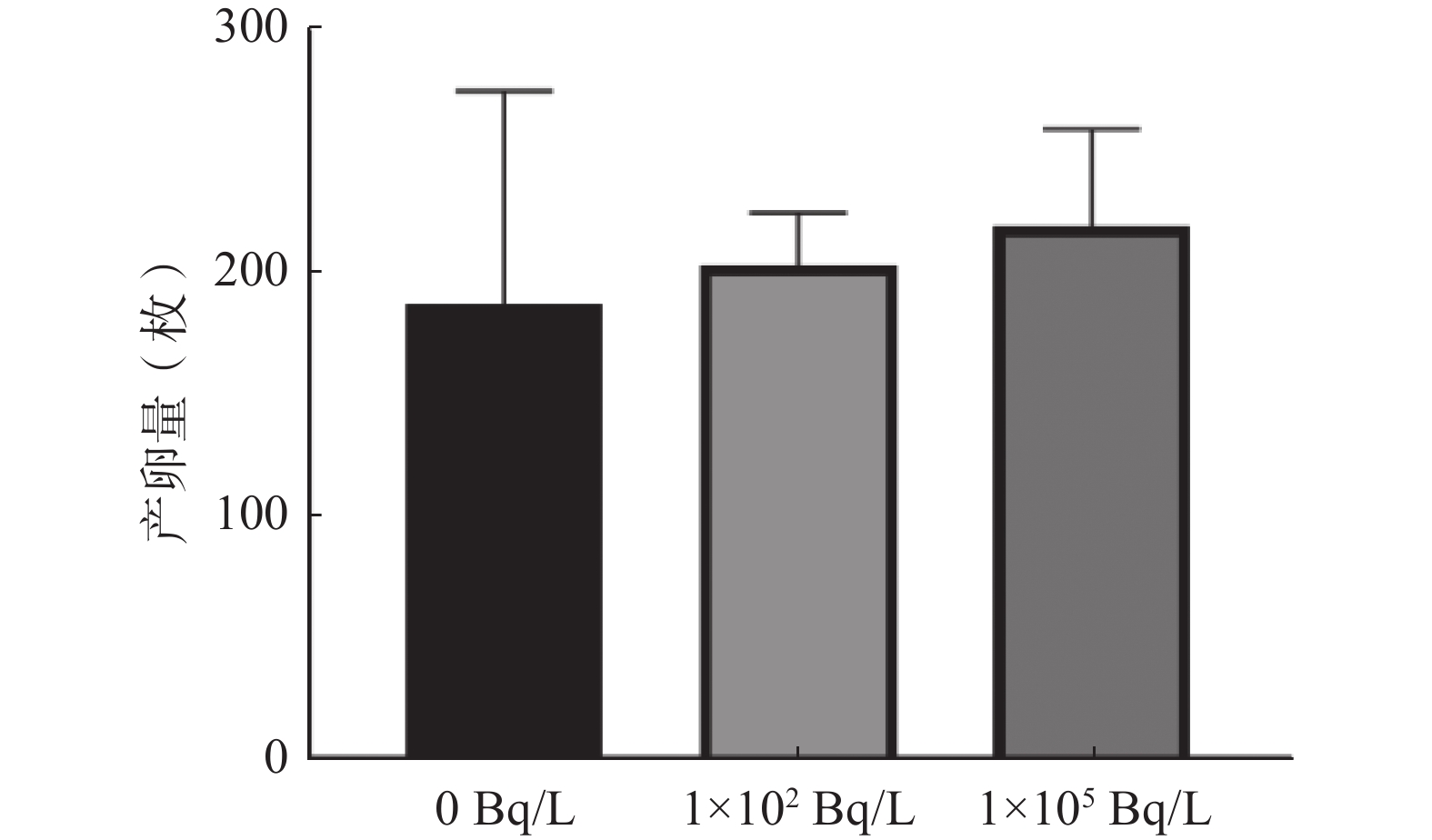
由图5可知,0、1×102、1×105 Bq/L氚水暴露组斑马鱼产卵量分别为(188±88)、(204±22)、(220±40)枚。与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼产卵量的差异均无统计学意义(t=0.400、0.757,P=0.700、0.477)。

图 5 不同浓度氚水长期暴露F1代斑马鱼成鱼产卵量的比较
Figure 5. Comparison of eggs laid by adult F1 generation zebrafish after long-term exposure to different concentrations of tritiated water
-
1×105 Bq/L氚水长期暴露条件下,F1代斑马鱼的鱼龄达60 d时,测得其体内总氚含量为(32.23±1.97) Bq/g。
-
在研究低剂量氚水长期暴露所致生物效应时需要选取合适的暴露浓度与实验动物模型。目前对于氚水浓度高低划分的标准尚未统一,且不同国家之间差异显著。以饮用水中氚含量的限值为例,欧盟大部分国家的标准为1×102 Bq/L,加拿大为7×103 Bq/L,俄罗斯为7.7×103 Bq/L,芬兰为3×104 Bq/L,澳大利亚为>7.6×103 Bq/L[13]。同时,核电站排放的氚水浓度通常为1×106 Bq/L[14]。对于在内陆建造的核电站,我国要求其排放口下游1 km处受纳水体中氚浓度不超过1×102 Bq/L[15]。因此,本文综合环境因素选取低剂量氚水暴露浓度为1×102和1×105 Bq/L。
由于低剂量氚水的生物效应不易被观察到,一些研究者通过构建更为敏感的动物模型,如转入Rev1基因的C57BL/6N小鼠、转染人源X染色体的Hprt基因缺失的仓鼠细胞等对低剂量氚水的生物效应进行研究[16-17]。然而这些模型目前并不适用于低剂量氚水长期暴露的生物效应研究。斑马鱼作为一种理想的实验动物模型被广泛应用于氚水的毒性效应研究中,且由于其体型小、养成周期短、产卵量大、胚胎透明、成本低等优势十分适合用于氚水长期暴露的生物效应研究[18-19]。Li等[20]研究发现,氚水对斑马鱼胚胎的行为、生理和基因的表达产生综合影响。Arcanjo等[21-22]研究发现,氚水暴露后斑马鱼幼苗参与肌肉收缩、眼晶体透明度、DNA损伤修复的基因出现错误表达。虽然上述研究中的氚水浓度均高于本实验设置的浓度,但是考虑斑马鱼作为模型动物的优势,可以对其在低浓度氚水暴露方面的研究进行探索。
本研究中,在低剂量氚水长期暴露的条件下,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼的孵化率、自主运动、心率、体长、产卵量等指标的差异均无统计学意义,说明斑马鱼在1×102、1×105 Bq/L浓度的氚水中仅暴露到F1代并未对其基本生长发育造成影响。目前,氚水长期暴露的生物效应的相关研究较少,一些发现氚水暴露对斑马鱼生长发育指标产生影响的研究中所设置的氚水浓度较高且均为急性暴露[20,23],因此本实验结果仍需做进一步研究。
氧化应激相关指标(ROS、SOD、MDA等)在氚水的毒性效应评价中得到了广泛应用,但不同浓度氚水暴露的生物效应仍不明确[12]。Huang等[24]对在3.7×106 Bq/L氚水中暴露96 h的小球藻和莱茵衣藻的氧化应激相关指标进行检测,结果表明,小球藻的ROS荧光强度增加、SOD含量降低,MDA含量无明显变化;莱茵衣藻的ROS荧光强度、SOD含量、MDA含量均无明显变化,与之前的研究结果[25]不一致,他们认为可能是由于氚水的暴露浓度与时间不一致所致。Gagnaire等[26]对在1×108 Bq/L氚水中暴露的斑马鱼进行了ROS检测,结果表明,鱼龄为4 d的斑马鱼的ROS荧光强度增加,但是鱼龄为7 d和10 d的斑马鱼的ROS荧光强度无明显变化。本研究中,与0 Bq/L氚水暴露组比较,1×102 Bq/L氚水暴露组斑马鱼的ROS荧光强度、T-SOD含量、MDA含量的差异均无统计学意义;1×105 Bq/L氚水暴露组60 d的 F1代斑马鱼的MDA含量降低。因此仍需就相关指标做进一步研究。
1×105 Bq/L氚水暴露组60 d 的F1代斑马鱼体内的总氚含量为(32.23±1.97) Bq/g。其他浓度氚水和暴露时间的样品未检测到明显的氚含量积累,其原因可能为样本中的氚含量过低或低本底液闪计数器的检测性能不足所致。斑马鱼体内氚含量的明显积累提示氚水长期暴露可能会对斑马鱼产生一定影响,较多研究结果均表明相同氚含量下有机结合氚比游离氚产生的生物效应更加显著[23, 27-29]。后续需要对氚水的暴露时间、暴露浓度、测量指标等做进一步优化,以更好地观察氚水长期暴露的毒性。
本研究依据核电站实际排放核废水的标准及环境水体中氚浓度的变化设置了氚水浓度,并以斑马鱼为实验模型对该浓度氚水长期暴露所致F1代斑马鱼的生物效应做了初步探讨。未来应对氚水长期暴露斑马鱼子代作进一步研究,观察F1代后续子代的各项指标以进一步明确低剂量氚水长期暴露的生物效应,为环境评价和生物健康提供完善的参考数据。
利益冲突 所有作者声明无利益冲突
作者贡献声明 顾鹏诚负责现场实验的实施、论文的撰写;罗发坚、薛惠元负责实验模型的建立、数据的收集与分析;陈娜、孙亮、万骏负责实验思路的提出与实验设计的指导;崔凤梅、涂彧负责论文的审阅与修改
氚水长期暴露对斑马鱼子代生长发育影响的研究
Effects of long-term exposure to tritiated water on the growth and development of zebrafish offspring
-
摘要:
目的 研究氚水长期暴露对斑马鱼子代生长发育的影响。 方法 将野生型AB品系斑马鱼所产胚胎暴露在0、1×102、1×105 Bq/L氚水中长期饲养作为亲代(F0代),待其性成熟后进行繁殖,所得子代作为F1代。F1代斑马鱼继续饲养在与F0代对应浓度的氚水中。观察F1代斑马鱼的生长发育情况,检测胚胎期的自主运动、心率,幼苗期的孵化率、体长、活性氧(ROS)荧光强度,幼鱼期的总超氧化物歧化酶(T-SOD)、丙二醛(MDA)、总氚含量,成鱼期的产卵量。各检测指标的组间比较采用t检验(方差齐)。 结果 F1代0、1×102、1×105 Bq/L氚水暴露组斑马鱼的孵化率分别为(90.66±0.05)%、(85.63±0.10)%、(78.06±0.15)%,与0 Bq/L氚水暴露组相比, 1×102、1×105 Bq/L氚水暴露组F1代斑马鱼孵化率的差异均无统计学意义(t=0.785、1.370,P=0.462、0.220)。F1代3组斑马鱼受精后24 h的自主运动次数分别为(12.93±2.70)、(11.30±0.78)、(10.50±0.80) 次/min,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后24 h自主运动次数的差异均无统计学意义(t=1.008、1.499,P=0.370、0.208)。3组斑马鱼受精后36 h的自主运动次数分别为(3.63±1.43)、(4.50±1.15)、(5.40±3.55) 次/min,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后36 h自主运动次数的差异均无统计学意义(t=0.817、0.799,P=0.460、0.469)。3组斑马鱼受精后48 h的心率分别为(59.43±6.93)、(65.00±3.30)、(61.23±4.55) 次/20 s,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后48 h心率的差异均无统计学意义(t=1.256、0.376,P=0.278、0.726)。3组斑马鱼受精后60 h的心率分别为(69.87±2.71)、(66.17±6.97)、(69.77±9.08) 次/20 s,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后60 h心率的差异均无统计学意义(t=0.857、0.018,P=0.440、0.986)。3组斑马鱼受精后72 h的体长分别为(3.20±0.22)、(3.32±0.08)、(3.29±0.06) mm,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后72 h体长的差异均无统计学意义(t=0.614、0.178,P=0.525、0.868)。3组斑马鱼受精后84 h的体长分别为(3.42±0.07)、(3.46±0.11)、(3.40±0.04) mm,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼受精后84 h体长的差异均无统计学意义(t=0.527、0.496,P=0.626、0.646)。F1代3组斑马鱼ROS荧光强度分别为(21.07±4.74)、(23.71±7.73)、(23.19±5.32),与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼幼苗ROS荧光强度的差异均无统计学意义(t=0.582、0.593,P=0.582、0.575)。F1代3组斑马鱼在45 d时的T-SOD含量分别为(41.84±4.91)、(42.30±5.04)、(36.97±5.26) U/mgprot,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼45 d T-SOD含量的差异均无统计学意义(t=0.112、1.171,P=0.916、0.307)。3组斑马鱼60 d的T-SOD含量分别为(36.93±1.91)、(34.07±3.02)、(33.54±1.87) U/mgprot,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼45 d T-SOD含量的差异均无统计学意义(t=1.397、2.195,P=0.240、0.093)。3组斑马鱼45 d的MDA含量分别为(3.60±1.56)、(3.59±0.44)、(2.95±0.58) nmol/mgprot,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼MDA含量的差异均无统计学意义(t=0.007、0.677,P=0.995、0.536)。3组斑马鱼60 d的MDA含量分别为(4.00±0.52)、(4.19±1.37)、(3.01±0.32) nmol/mgprot,与0 Bq/L氚水暴露组相比,1×102 Bq/L氚水暴露组F1代斑马鱼60 d MDA含量的差异无统计学意义(t=0.229,P=0.830),1×105 Bq/L氚水暴露组F1代斑马鱼60 d MDA含量的差异有统计学意义(t=2.831,P=0.047)。F1代3组斑马鱼产卵量分别为(188±88)、(204±22)、(220±40)枚,与0 Bq/L氚水暴露组相比,1×102、1×105 Bq/L氚水暴露组F1代斑马鱼产卵量的差异均无统计学意义(t=0.400、0.757,P=0.700、0.477)。1×105 Bq/L 氚水长期暴露可导致F1代斑马鱼体内总氚含量的累积,鱼龄60 d时其体内总氚含量为(32.23±1.97) Bq/g。 结论 1×105 Bq/L氚水长期暴露可导致F1代斑马鱼体内的氚含量积累。 Abstract:Objective To study the effects of long-term tritiated water exposure on the growth and development of zebrafish offspring. Methods Embryos produced by wild-type AB strain zebrafish were exposed to 0, 1×102, and 1×105 Bq/L tritiated water for long-term feeding as parents (F0 generation). After their sexual maturity, they reproduced, and the offspring obtained were recorded as F1 generation. The F1 generation zebrafish continued to be raised in tritiated water concentrations corresponding to the F0 generation. We observed the growth and development of F1 generation zebrafish and detected autonomous movement and heart rate during the embryonic stage; hatching rate, body length, and reactive oxygen species (ROS) fluorescence intensity during the seedling stage; total superoxide dismutase (T-SOD), malondialdehyde (MDA), and total tritium contents during the juvenile stage; and egg production during the adult stage. The t-test was used for intergroup comparison of various detection indicators(equal variance). Results The hatching rates of the three groups of zebrafish in F1 generation were (90.66±0.05)%, (85.63±0.10)%, and (78.06±0.15)%. Compared with the 0 Bq/L tritiated water exposure group, no statistically significant difference was found in the hatching rate of F1 zebrafish between the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.785, 1.370; P=0.462, 0.220). The number of autonomous movement of the three groups of zebrafish in F1 generation at 24 h after fertilization was (12.93±2.70), (11.30±0.78), and (10.50±0.80) times/min. Compared with the 0 Bq/L tritiated water exposure group, we observed no statistically significant difference in the number of autonomous movements of F1 generation zebrafish at 24 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=1.008, 1.499; P=0.370, 0.208). The number of autonomous movement of the three groups of zebrafish at 36 h after fertilization was (3.63±1.43), (4.50±1.15), and (5.40±3.55) times/min. Compared with the 0 Bq/L tritiated water exposure group, we found no statistically significant difference in the number of autonomous movement of F1 generation zebrafish at 36 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.817, 0.799; P=0.460, 0.469). The heart rates of the three groups of zebrafish at 48 h after fertilization were (59.43±6.93), (65.00±3.30), and (61.23±4.55) times/20 s. Compared with the 0 Bq/L tritiated water exposure group, we observed no statistically significant difference in the heart rate of F1 generation zebrafish at 48 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=1.256, 0.376; P=0.278, 0.726). The heart rates of the three groups of zebrafish at 60 h after fertilization were (69.87±2.71), (66.17±6.97), and (69.77±9.08) times/20 s. Compared with the 0 Bq/L tritiated water exposure group, we found no statistically significant difference in the heart rate of F1 generation zebrafish at 60 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.857, 0.018; P=0.440, 0.986).The body lengths of the three groups of zebrafish at 72 h after fertilization were (3.20±0.22), (3.32±0.08), and (3.29±0.06) mm. Compared with the 0 Bq/L tritiated water exposure group, no statistically significant difference was noted in the body length of F1 generation zebrafish at 72 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.614, 0.178; P=0.525, 0.868). The body length of the three groups of zebrafish at 84 h after fertilization were (3.42±0.07), (3.46±0.11), and (3.40±0.04) mm. Compared with the 0 Bq/L tritiated water exposure group, no statistically significant difference was observed in the body length of F1 generation zebrafish at 84 h after fertilization in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.527, 0.496; P=0.626, 0.646). The ROS fluorescence intensities of the three groups of zebrafish in F1 generation were (21.07±4.74), (23.71±7.73), and (23.19±5.32), respectively. Compared with the 0 Bq/L tritiated water exposure group, there was no statistically significant difference in ROS fluorescence intensity of F1 generation zebrafish seedlings in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.582, 0.593; P=0.582, 0.575). The T-SOD contents of the three groups of zebrafish in F1 generation at 45 days were (41.84±4.91), (42.30±5.04), and (36.97±5.26) U/mgprot. Compared with the 0 Bq/L tritiated water exposure group, we found no statistically significant difference in the T-SOD content of F1 generation zebrafish at 45 days in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.112, 1.171; P=0.916, 0.307).The T-SOD contents of three groups of zebrafish at 60 days were (36.93±1.91), (34.07±3.02), and (33.54±1.87) U/mgprot. Compared with the 0 Bq/L tritiated water exposure group, there was no statistically significant difference in the T-SOD content of F1 generation zebrafish at 60 days in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=1.397, 2.195; P=0.240, 0.093). The MDA contents of the three groups of zebrafish at 45 days were (3.60±1.56), (3.59±0.44), and (2.95±0.58) nmol/mgprot. Compared with the 0 Bq/L tritiated water exposure group, we found no statistically significant difference in the MDA content of F1 generation zebrafish at 45 days in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.007, 0.677; P=0.995, 0.536). The MDA contents of the three groups of zebrafish at 60 days were (4.00±0.52), (4.19±1.37), and (3.01±0.32) nmol/mgprot. Compared with the 0 Bq/L tritiated water exposure group, there was no statistically significant difference in the MDA content in F1 generation zebrafish at 60 days in the 1×102 Bq/L tritiated water (t=0.229, P=0.830); the difference in the MDA content in F1 generation zebrafish exposed to 1×105 Bq/L tritiated water was statistically significant (t=2.831, P=0.047). The eggs laid by the three groups of zebrafish in F1 generation were 188±88, 204±22, and 220±40. Compared with the 0 Bq/L tritiated water exposure group, we found no statistically significant difference in the egg production of F1 generation zebrafish in the 1×102 Bq/L and 1×105 Bq/L tritiated water exposure groups (t=0.400, 0.757; P=0.700, 0.477). The tritium content in the body of F1 generation zebrafish was (32.23±1.97) Bq/g at 60 days. Conclusion Long-term exposure to 1×105 Bq/L tritiated water can lead to the accumulation of tritium in F1 generation zebrafish. -
Key words:
- Tritium /
- Zebrafish /
- F1 generation /
- Biological effects
-
图 1 不同浓度氚水长期暴露F1代斑马鱼孵化率的比较
Figure 1. Comparison of hatching rate of F1 generation zebrafish after long-term exposure to different concentrations of tritiated water
图 2 不同浓度氚水长期暴露F1代斑马鱼受精后不同时间自主运动次数、心率、体长的比较
Figure 2. Comparison of autonomous movement, heart rate and body length of F1 generation zebrafish at different times after fertilization after long-term exposure to different concentrations of tritiated water
图 3 不同浓度氚水长期暴露F1代斑马鱼幼苗活性氧荧光强度的比较
Figure 3. Comparison of reactive oxygen species fluorescence intensity of F1 generation zebrafish seedlings after long-term exposure to different concentrations of tritiated water
图 4 不同浓度氚水长期暴露F1代不同鱼龄斑马鱼幼鱼体内T-SOD、MDA含量的比较
Figure 4. Comparison of total superoxide dismutase and malondialdehyde content in F1 generation zebrafish of different age after long-term exposure to different concentrations of tritiated water
-
[1] Galeriu D, Heling R, Melintescu A. The dynamics of tritium - including OBT - in the aquatic food chain[J]. Fusion Sci Technol, 2005, 48(1): 779−782. DOI: 10.13182/fst05-a1036. [2] Yamanishi T, Kakiuchi H, Tauchi H, et al. Discussions on tritiated water treatment for Fukushima Daiichi Nuclear Power Station[J]. Fusion Sci Technol, 2020, 76(4): 430−438. DOI: 10.1080/15361055.2020.1716454. [3] Zhao C, Wang G, Zhang M, et al. Transport and dispersion of tritium from the radioactive water of the Fukushima Daiichi nuclear plant[J]. Mar Pollut Bull, 2021, 169: 112515. DOI: 10.1016/j.marpolbul.2021.112515. [4] 张琪轩. 氚水和木黄酮联合染毒对斑马鱼生长发育的毒性作用及机制研究[D]. 苏州: 苏州大学, 2022. DOI: 10.27351/d.cnki.gszhu.2022.000076.
Zhang QX. Toxicity and mechanism of tritium water and genistein combined exposure to growth and development of zebrafish[D]. Suzhou: Soochow University, 2022. DOI: 10.27351/d.cnki.gszhu.2022.000076.[5] Mountford PJ, Temperton DH. Recommendations of the International Commission on Radiological Protection (ICRP) 1990[J]. Eur J Nucl Med, 1992, 19(2): 77−79. DOI: 10.1007/BF00184120. [6] Little MP, Lambert BE. Systematic review of experimental studies on the relative biological effectiveness of tritium[J]. Radiat Environ Biophys, 2008, 47(1): 71−93. DOI: 10.1007/s00411-007-0143-y. [7] Siragusa M, Fredericia PM, Jensen M, et al. Radiobiological effects of tritiated water short-term exposure on V79 clonogenic cell survival[J]. Int J Radiat Biol, 2018, 94(2): 157−165. DOI: 10.1080/09553002.2018.1419301. [8] 彭述明, 周晓松, 陈志林. 氚化学与氚分析进展与展望[J]. 核化学与放射化学, 2020, 42(6): 498−512. DOI: 10.7538/hhx.2020.YX.2020075.
Peng SM, Zhou XS, Chen ZL. Progress of tritium chemistry and tritium analysis[J]. J Nucl Radiochem, 2020, 42(6): 498−512. DOI: 10.7538/hhx.2020.YX.2020075.[9] 李圣日. 氚水对斑马鱼生长发育的毒性作用及机制研究[D]. 苏州: 苏州大学, 2021. DOI: 10.27351/d.cnki.gszhu.2021.003315.
Li SR. Study on the toxicity and mechanism of tritiated water on the growth and development of zebrafish[D]. Suzhou: Soochow University, 2021. DOI: 10.27351/d.cnki.gs zhu.2021.003315.[10] Dang ZC, Traas T, Vermeire T. Evaluation of the fish short term reproduction assay for detecting endocrine disrupters[J]. Chemosphere, 2011, 85(10): 1592−1603. DOI: 10.1016/j.chemosphere.2011.08.006. [11] Constantine LA, Green JW, Schneider SZ. Ibuprofen: fish short-term reproduction assay with zebrafish ( Danio rerio) based on an extended OECD 229 protocol[J]. Environ Toxicol Chem, 2020, 39(8): 1534−1545. DOI: 10.1002/etc.4742. [12] Kelsey-Wall A, Seaman JC, Jagoe CH, et al. Biological half-life and oxidative stress effects in mice with low-level, oral exposure to tritium[J]. J Toxicol Environ Health A, 2006, 69(3): 201−213. DOI: 10.1080/15287390500227365. [13] Guéguen Y, Priest ND, Dublineau I, et al. In vivo animal studies help achieve international consensus on standards and guidelines for health risk estimates for chronic exposure to low levels of tritium in drinking water[J]. Environ Mol Mutagen, 2018, 59(7): 586−594. DOI: 10.1002/em.22200. [14] Jaeschke BC, Bradshaw C. Bioaccumulation of tritiated water in phytoplankton and trophic transfer of organically bound tritium to the blue mussel, Mytilus edulis[J]. J Environ Radioact, 2013, 115: 28−33. DOI: 10.1016/j.jenvrad.2012.07.008. [15] 张姝雅, 杨钱, 金爱平, 等. 水精馏技术在氚化水分离中的应用进展[J]. 核技术, 2023, 46(6): 25−38. DOI: 10.11889/j.0253-3219.2023.hjs.46.060301.
Zhang SY, Yang Q, Jin AP, et al. Progress in the application of water distillation for tritiated water separation[J]. Nucl Tech, 2023, 46(6): 25−38. DOI: 10.11889/j.0253-3219.2023.hjs.46.060301.[16] Tauchi H, Imamura H, Inoue M, et al. Assessment of biological effect of tritiated water by using hypersensitive system[J]. Fusion Sci Technol, 2011, 60(3): 1173−1178. DOI: 10.13182/fst11-a12624. [17] Toyoshima M, Honda H, Watanabe H, et al. Development of a sensitive assay system for tritium risk assessment using Rev1 transgenic mouse[J]. Fusion Sci Technol, 2011, 60(3): 1204−1207. DOI: 10.13182/fst11-a12632. [18] Canedo A, Rocha TL. Zebrafish ( Danio rerio) using as model for genotoxicity and DNA repair assessments: historical review, current status and trends[J]. Sci Total Environ, 2021, 762: 144084. DOI: 10.1016/j.scitotenv.2020.144084. [19] Gagnaire B, Arcanjo C, Cavalié I, et al. Effects of gamma ionizing radiation exposure on Danio rerio embryo-larval stages - comparison with tritium exposure[J]. J Hazard Mater, 2021, 408: 124866. DOI: 10.1016/j.jhazmat.2020.124866. [20] Li SR, Zhang YF, Xue HY, et al. Integrative effects based on behavior, physiology and gene expression of tritiated water on zebrafish[J]. Ecotox Environ Safe, 2021, 225: 112770. DOI: 10.1016/j.ecoenv.2021.112770. [21] Arcanjo C, Armant O, Floriani M, et al. Tritiated water exposure disrupts myofibril structure and induces mis-regulation of eye opacity and DNA repair genes in zebrafish early life stages[J]. Aquat Toxicol, 2018, 200: 114−126. DOI: 10.1016/j.aquatox.2018.04.012. [22] Arcanjo C, Adam-Guillermin C, Murat El Houdigui S, et al. Effects of tritiated water on locomotion of zebrafish larvae: a new insight in tritium toxic effects on a vertebrate model species[J]. Aquat Toxicol, 2020, 219: 105384. DOI: 10.1016/j.aquatox.2019.105384. [23] Quan Y, Tan ZY, Yang Y, et al. Prolonged effect associated with inflammatory response observed after exposure to low dose of tritium β-rays[J]. Int J Radiat Biol, 2020, 96(8): 972−979. DOI: 10.1080/09553002.2020.1767817. [24] Huang Y, Qin M, Lai JL, et al. Assessing OBT formation and enrichment: ROS signaling is involved in the radiation hormesis induced by tritium exposure in algae[J]. J Hazard Mater, 2023, 443: 130159. DOI: 10.1016/j.jhazmat.2022.130159. [25] Réty C, Gilbin R, Gomez E. Induction of reactive oxygen species and algal growth inhibition by tritiated water with or without copper[J]. Environ Toxicol, 2012, 27(3): 155−165. DOI: 10.1002/tox.20626. [26] Gagnaire B, Arcanjo C, Cavalié I, et al. Tritiated water exposure in zebrafish ( Danio rerio): effects on the early-life stages[J]. Environ Toxicol Chem, 2020, 39(3): 648−658. DOI: 10.1002/etc.4650. [27] Roch-Lefèvre S, Grégoire E, Martin-Bodiot C, et al. Cytogenetic damage analysis in mice chronically exposed to low-dose internal tritium beta-particle radiation[J/OL]. Oncotarget, 2018, 9(44): 27397−27411[2023-08-28]. https://www.oncotarget.com/article/25282/text. DOI: 10.18632/oncotarget.25282. [28] Galeriu D, Davis P, Raskob W, et al. Recent progresses in tritium radioecology and dosimetry[J]. Fusion Sci Technol, 2008, 54(1): 237−242. DOI: 10.13182/fst08-a1803. [29] Kim SB, Baglan N, Davis PA. Current understanding of organically bound tritium (OBT) in the environment[J]. J Environ Radioact, 2013, 126: 83−91. DOI: 10.1016/j.jenvrad.2013.07.011. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1405
- HTML全文浏览量: 1309
- PDF下载量: 11
