


 下载:
下载:
-
放射疗法是胸部肿瘤的治疗方法之一,其通常会引起放射性肺损伤(radiation-induced lung injury,RILI),发生率为5%~20%[1]。RILI是一个复杂的动态过程,涉及多种细胞、途径和机制[2-3]。有研究结果显示,胸部在受到照射后的早期,肺组织会出现明显的免疫失衡现象[4-5]。我们的前期研究结果显示,辐射损伤的肺上皮细胞可能会刺激树突状细胞的抗原呈递功能进而激活T细胞;在树突状细胞不存在的情况下,辐射损伤的肺上皮细胞依旧能活化T细胞[6]。外泌体是具有双层膜结构的细胞外囊泡,其可以携带信息物质参与细胞间通信。Ikhlas等[7]的研究结果显示,受损的非专职性抗原呈递细胞分泌的外泌体可以参与免疫反应。而Liu等[8]的研究结果显示,肺组织中的细胞外囊泡主要来源于Ⅱ型肺上皮细胞。我们通过体外模拟肺部微环境中的细胞间通信,以期发现外泌体参与RILI过程的途径,为RILI的临床研究提供新的实验依据。
-
小鼠肺上皮MLE-12细胞(简称MLE-12细胞)购自美国模式培养物集存库(ATCC)细胞库。无特定病原体级雄性C57BL/6 WT小鼠购自北京维通利华实验动物有限公司[动物许可证号:SCXK(京)2021-0006],6~8周龄、体重(20±2) g。所有动物实验均在军事科学院军事医学研究院实验动物管理与使用委员会的指导下进行,本研究经军事科学院军事医学研究院伦理委员会批准(批准文号:IACUC-DWZX-2021-731)。胎牛血清(fetal bovine serum,FBS)购自中国四季青公司;RPMI-1640培养基购自美国Gibco公司;外泌体流式试剂盒购自日本Wako公司;EasySep小鼠初始T细胞分离试剂盒购自加拿大Stemcell公司;外泌体抑制剂GW4869购自美国MedChemExpress公司;细胞蛋白裂解液和增强型化学发光试剂(ECL)发光液购自美国Thermo公司;外泌体专用裂解液购自中国宇玫博公司;BCA蛋白质浓度测定试剂盒和牛血清白蛋白(简称BSA)购自中国翌圣公司;鼠抗鼠肿瘤易感基因(tumor susceptibility gene,TSG)101、溶酶体相关膜蛋白3[别称分化抗原簇(cluster of differentiation,CD)63]和四次跨膜蛋白28(别称CD81)抗体购自美国Santacruz公司;兔抗鼠Ⅰ型内质网膜蛋白(Calnexin)抗体购自中国爱博泰克公司;兔抗鼠Ⅰ类主要组织相容性复合体(major histocompatibility complex class Ⅰ,MHC Ⅰ)抗体和兔抗鼠Ⅱ类主要组织相容性复合体 (major histocompatibility complex class Ⅱ,MHC Ⅱ)抗体购自中国碧云天公司;山羊抗兔免疫球蛋白(immunoglobulin,Ig)G(H+L)-辣根过氧化物酶(HRP)和山羊抗小鼠IgG(H+L)-HRP购自中国三箭公司;抗小鼠MHC Ⅰ-AF647、早期活化抗原1(别称CD69)-BV421抗体和死细胞染料购自美国Biolegend公司;抗小鼠MHC Ⅱ-异硫氰酸荧光素(FITC)、免疫调节蛋白B7-1(别称CD80)-PerCP-Cy5.5、免疫调节蛋白B7-2(别称CD86)-PE-Cy7、CD3-FITC、CD4-PE-Cy7、CD8-PerCP和T细胞特定表面糖蛋白CD28-PE抗体购自美国BD公司。FlowJo 10.6.2软件购自美国Tree Star公司;37℃恒温培养箱购自美国Thermo公司;CP100NX超速离心机购自日本Himac公司;Zetaview纳米颗粒跟踪分析仪购自德国PMX公司;H-7650透射电子显微镜(transmission electron microscope,TEM)购自日本HITACHI公司;BD FACS AriaTM Ⅱ流式细胞仪购自美国BD公司;R2089型60Co γ射线照射源购自英国维瑞斯公司,剂量率为59.09 cGy/min,位于军事科学院军事医学研究院60Co源照射室。
-
照射前12 h,将2×106个MLE-12细胞接种于含10% FBS的RPMI-1640培养基的培养皿中,培养条件为37℃、5%CO2,并将其分为对照组和γ射线照射组(2、4、6、8 Gy)。照射后用PBS洗涤细胞2次,将培养基改为含有去外泌体(简称D-exo)的10%FBS的RPMI-1640培养基。在4℃条件下,将FBS经100 000×g(P70AT转子)超速离心18 h,以去除FBS来源的外泌体,制为D-exo FBS。
D-exo FBS培养基培养MLE-12细胞12、24、48 h后收集其上清液以提取外泌体。上清液经300×g离心10 min去除细胞,经2 000×g离心10 min去除死细胞,经10 000×g离心30 min去除细胞碎片。用0.22 μm的过滤器过滤以上经过多步离心的培养基。过滤后的培养基经100 000×g(P70AT转子)超速离心1.5 h提取外泌体。弃上清,加入13 ml PBS洗涤外泌体,100 000×g(P40ST转子)条件下再次超速离心1.5 h沉淀外泌体。上述离心操作均在4℃条件下完成。将外泌体重悬于100 μl PBS(或适量蛋白裂解液)中,用于进一步分析或细胞共培养。
采用TEM和蛋白质印迹法(Western blot,WB)对外泌体进行鉴定。采用纳米颗粒跟踪分析技术(简称NTA)测量外泌体的粒径和数量。外泌体流式试剂盒用于间接分析外泌体表面标志物MHC Ⅰ、MHC Ⅱ、CD80和CD86。
-
使用EasySep小鼠初始T细胞分离试剂盒从雄性C57BL/6 WT小鼠脾脏的单细胞悬浮液中分离CD3+初始T细胞。分离细胞结束后,在含有10% FBS的RPMI-1640培养基中将T细胞浓度调节至1×106/ml。
为了探索外泌体是否激活T细胞,将初始T细胞分别与6 Gy γ射线(依据前期实验室构建的细胞共培养模型,我们选择γ射线照射的剂量为6 Gy[6])照射组的MLE-12细胞(简称IR MLE-12)产生的外泌体(简称exo/IR-MLE)和对照组MLE-12细胞(简称NC MLE-12)分泌的外泌体(简称exo/NC-MLE)共培养12、24 h,采用流式细胞术(flow cytometry,FCM)检测T细胞活化标志物CD28和CD69。
纳米颗粒跟踪分析技术检测外泌体浓度后,将其稀释至5×109/ml。将初始T细胞以1×106个/孔接种在6孔板中,并与外泌体以1∶500的比例分别共培养12、24 h。
-
为了探索抑制外泌体后的MLE-12细胞是否不再激活T细胞,将初始T细胞分别与IR MLE-12、NC MLE-12和外泌体抑制剂GW4869处理的MLE-12细胞共培养。6 Gy γ射线照射前12 h,将MLE-12细胞以4×105个/孔接种在6孔板中。照射后,将初始T细胞和MLE-12细胞以5∶2的比例共培养,于12、24 h后收集重悬的细胞。外泌体抑制剂GW4869处理组在接种MLE-12细胞时和共培养期间均给予GW4869,其浓度为10 μmol/L。
-
使用0.25%的胰蛋白酶消化并收集MLE-12细胞,经PBS洗涤2次后,在4℃条件下,300×g离心5 min。将适量的细胞蛋白裂解液加入细胞沉淀中并于冰上孵育15 min。孵育结束后120 000×g离心10 min以沉淀细胞碎片,并将上清液转移至新管中。使用外泌体专用裂解液从外泌体中提取蛋白质。使用BCA蛋白质浓度测定试剂盒检测提取的蛋白质浓度。
转膜结束后,用含0.5%牛血清白蛋白的TBST(Tris Buffered Saline with Tween 20)缓冲液室温(25℃)孵育膜1 h以封闭膜上的未结合位点。封闭结束后,4℃孵育一抗12 h。用于检测目标蛋白的一抗为TSG101(1∶500稀释)、CD63(1∶500稀释)、CD81(1∶500稀释)、Calnexin(1∶1 000稀释)、MHC Ⅰ(1∶500稀释)和MHC Ⅱ(1∶1 000稀释)。一抗孵育结束后,用TBST洗涤膜3次,每次10 min。室温下孵育二抗(1∶4 000稀释)1 h,二抗孵育结束后,用TBST缓冲液洗涤膜3次,每次10 min。使用增强型化学发光试剂(ECL)曝光条带,曝光时间在5 min内。
-
用抗小鼠MHC Ⅰ-AF647抗体、抗小鼠MHC Ⅱ-FITC抗体、CD80-PerCP-Cy5.5抗体和CD86-PE-Cy7抗体标记外泌体-免疫磁珠复合物悬液,以检测外泌体表面抗原呈递相关分子MHC Ⅰ、MHC Ⅱ、CD80和CD86;用CD3-FITC抗体、CD4-PE-Cy7抗体、CD8-PerCP抗体、CD28-PE抗体、CD69-BV421抗体和死细胞染料标记T细胞悬液,以检测T细胞表面标志物CD3、CD4、CD8、CD28和CD69。使用流式细胞仪分析细胞,FlowJo 10.6.2软件分析数据,基于死细胞染料排除死细胞。
-
应用SPSS 19.0软件对数据进行统计学分析。符合正态分布的计量资料以
$ \bar x\pm s $ 表示,2组间比较采用两独立样本t检验(方差齐),多组间比较采用方差分析法分析,组间两两比较采用Bonferroni调整法。P<0.05为差异有统计学意义。 -
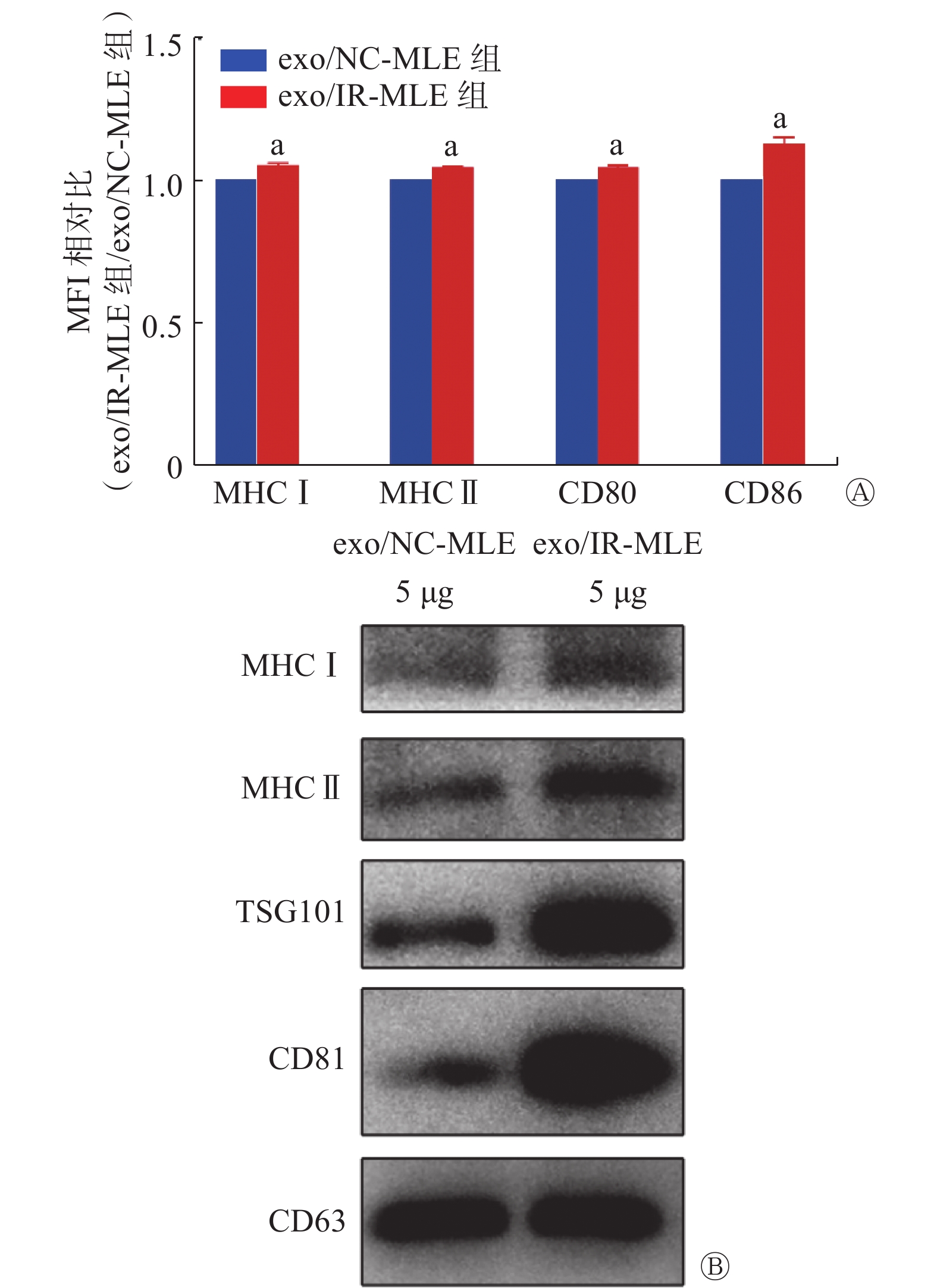
TEM和纳米颗粒跟踪分析技术结果显示,正常MLE-12细胞分泌的外泌体具有典型的一面凹陷的茶托样结构,粒径为30~150 nm(图1A、1B)。WB结果显示,与MLE-12细胞相比,其外泌体中特异性标志物CD63、CD81和TSG101高表达,而阴性标志物Calnexin低表达(图1C)。以上研究结果与国际细胞外囊泡协会发布的细胞外囊泡最小鉴定标准[9]一致。另外,与对照组相比,在6 Gy γ射线照射后不同时间(12、24和48 h)以及不同剂量γ射线(2、4、6和8 Gy)照射后24 h,单个MLE-12细胞分泌的外泌体数量于24、48 h时均明显增加(t=5.36、6.66,均P<0.05,图1D),且具有良好的剂量-效应关系,在照射剂量为6、8 Gy时差异有统计学意义(t=4.14、5.67,均P<0.05,图1E)。

图 1 γ射线照射前后小鼠肺上皮MLE-12细胞分泌的外泌体情况
Figure 1. Exosomes secreted by mouse lung epithelial MLE-12 cells before and after irradiated with γ ray
-
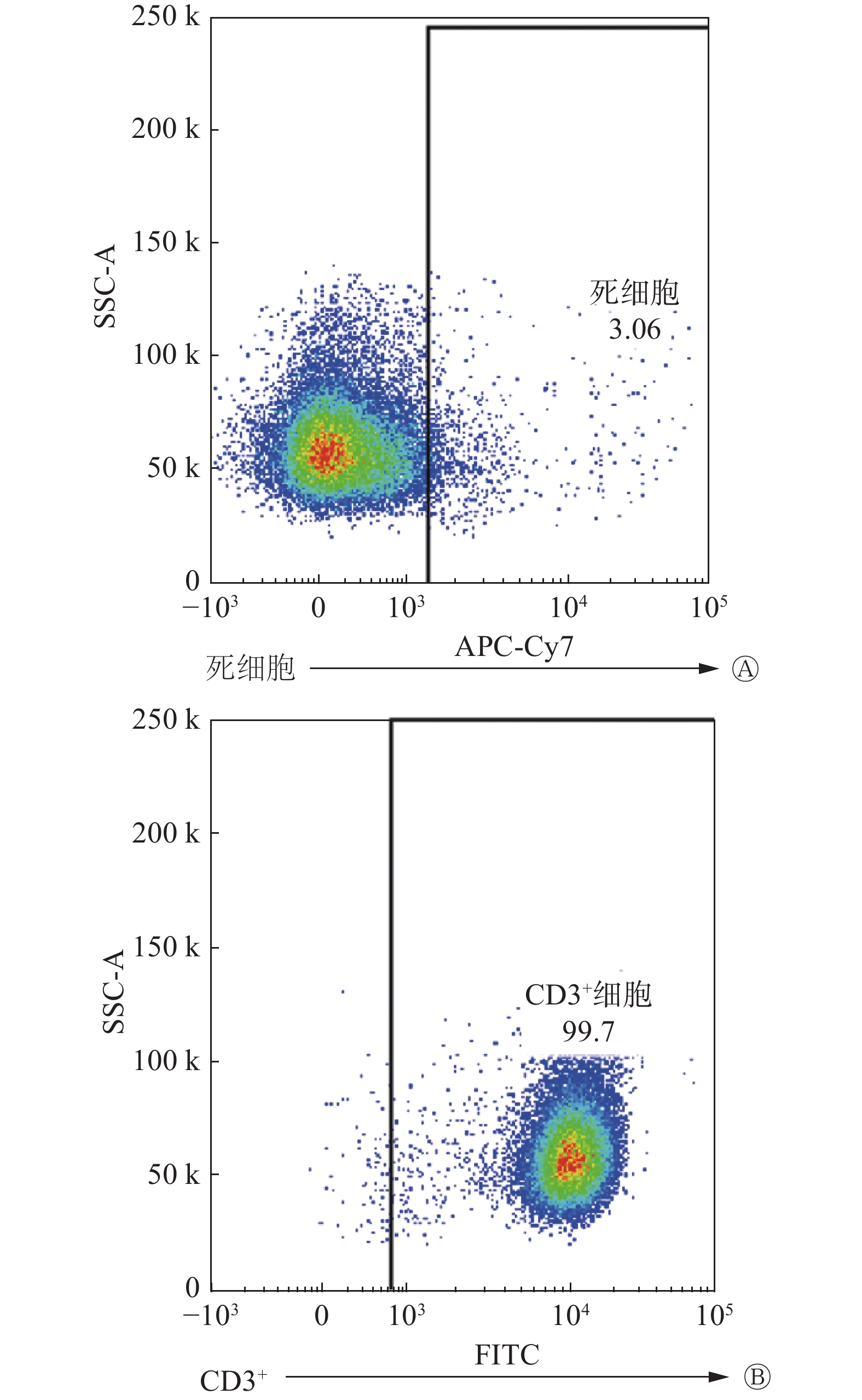
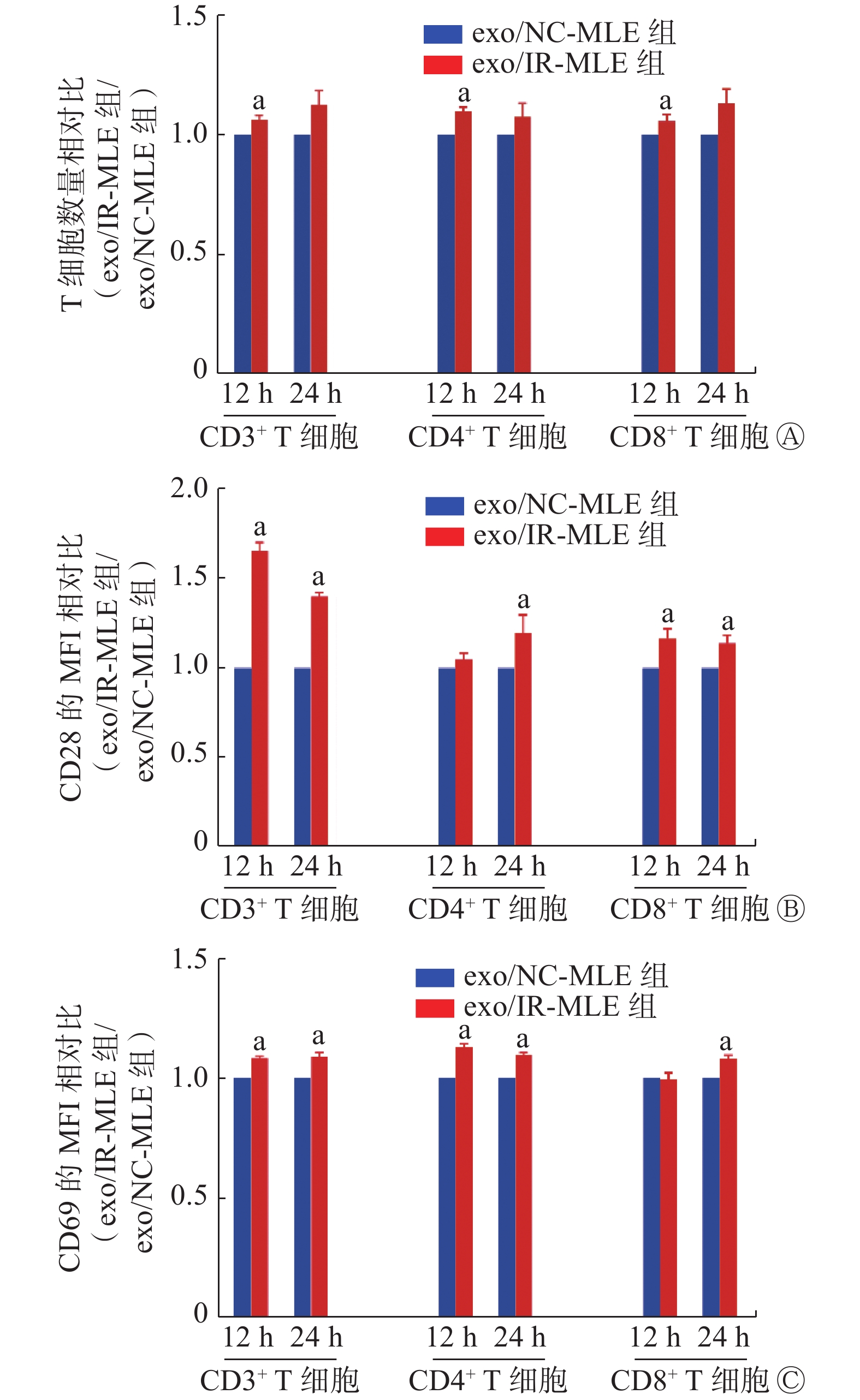
FCM结果显示,初始T细胞的纯度和活力均>95%(图2A、2B),符合实验需求。与exo/NC-MLE相比,exo/IR-MLE导致CD3+、CD4+和CD8+ T细胞数量增加(t=3.08~5.88,均P<0.05,图3A)。此外,在这些exo/IR-MLE诱导的T细胞及其亚群细胞中,CD28和CD69的表达水平均升高(t=3.02~8.65,均P<0.05,图3B、3C)。结果表明,照射后的MLE-12细胞分泌的外泌体可诱导初始T细胞增殖活化。

图 2 免疫磁珠分选初始T细胞的纯度和活力
Figure 2. Purity and vitality of naive T cells sorted by immunomagnetic beads

图 3 γ射线照射的小鼠肺上皮MLE-12细胞分泌的外泌体诱导T细胞活化
Figure 3. Activation of T cells by exosomes secreted by mouse lung epithelial MLE-12 cells irradiated with γ ray
-
FCM结果显示,与exo/NC-MLE相比,MHC Ⅰ、MHC Ⅱ、CD80和CD86在exo/IR-MLE中的表达水平均升高(t=4.04~6.47,均P<0.05,图4A);WB结果显示,与exo/NC-MLE相比,MHC Ⅰ和MHC Ⅱ在exo/IR-MLE中的表达水平均升高(图4B)。因此,受照射细胞的外泌体具有通过抗原呈递直接激活T细胞的能力。

图 4 抗原呈递相关分子在γ射线照射前后小鼠肺上皮MLE-12细胞分泌的外泌体中的表达
Figure 4. Expression of antigen presentation related molecules in exosomes secreted by mouse lung epithelial MLE-12 cells before and after irradiated with γ ray
-
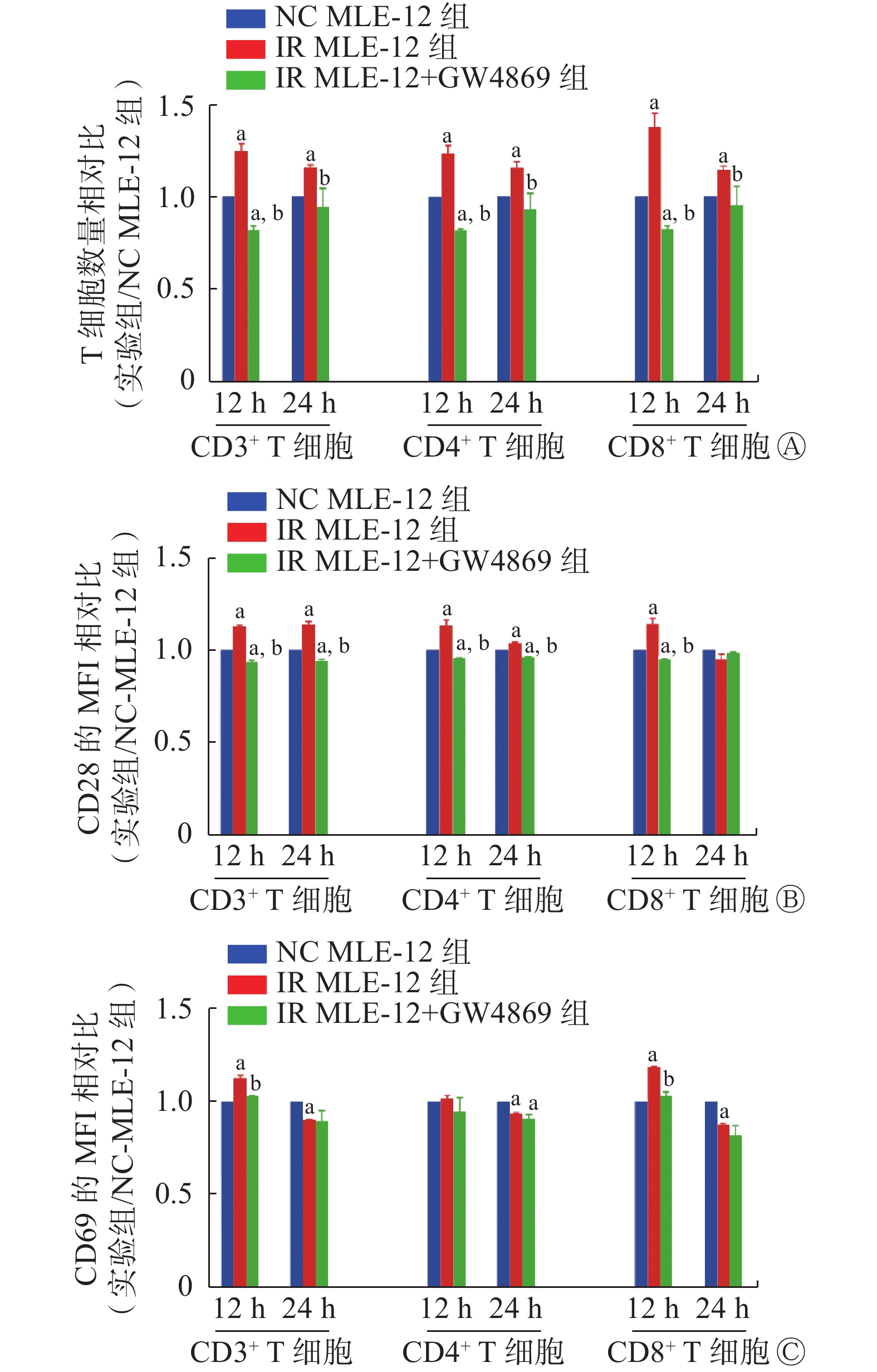
外泌体抑制剂GW4869抑制MLE-12细胞分泌外泌体的能力后,结果显示,IR MLE-12引起的CD3+、CD4+和CD8+ T细胞的数量增加以及CD28和CD69的表达增加(t=1.51~15.66,均P<0.05,图5)的现象均被明显抑制(t=3.64~23.03,均P<0.05),几乎与NC MLE-12细胞处理后的T细胞水平相同。上述研究结果显示,照射后的MLE-12细胞通过外泌体的抗原呈递激活T细胞。

图 5 外泌体抑制剂GW4869抑制γ射线照射后的小鼠肺上皮MLE-12细胞诱导的T细胞活化
Figure 5. T cell activation by mouse lung epithelial cells (MLE-12) irradiated with γ ray is suppressed by exosome inhibitor GW4869
-
本研究结果显示,照射后的MLE-12细胞分泌的外泌体可以诱导T细胞增殖活化,且T细胞表面CD28和CD69的表达水平均升高。由于CD28在T细胞活化期间接收第2信号参与抗原呈递[10],因此我们推测该活化T细胞的过程可能与抗原呈递有关。早期激活指标CD69的表达水平在12 h时升高,但在24 h时降低。Reddy等[11]的研究结果显示,T细胞活化后,CD69的表达水平首先随着时间的延长而升高,然后出现了显著降低的现象,原因是CD69的信号随着细胞增殖而被稀释[12],与本研究结果一致,即T细胞在12 h时被激活。上述研究结果表明,照射后的肺上皮细胞可以直接影响T细胞的活化,这与Berg等[13]报道的研究结果一致,其证实了RILI患者肺组织中存在T细胞活化的现象,即机体受到辐射后产生的应激反应会引起免疫反应[14]。因此T细胞活化参与RILI的进程,这可能是减缓RILI进展的关键潜在治疗靶标。
在共培养实验中,exo/IR-MLE激活T细胞并使其增殖。给予外泌体抑制剂GW4869后,T细胞活化现象受到抑制。同时,我们检测了一些抗原呈递相关分子在外泌体中的变化。与exo/NC-MLE相比,exo/IR-MLE中抗原呈递相关分子MHC Ⅰ、MHC Ⅱ、CD80和CD86表达水平均升高。结果显示,照射后的MLE-12细胞可以通过其分泌的外泌体激活T细胞,该激活过程可由抗原呈递介导。这一结果与Kulshreshtha等[15]的研究结果相似,他们证明了气道上皮细胞分泌的外泌体可以驱动单核细胞增殖,并增强其趋化性。Admyre等[16]的研究结果显示,人类肺泡灌洗液中的外泌体上存在共刺激分子,表明其具有直接调节T细胞活化的能力。上述研究结果为研究T细胞参与RILI的机制提供了新的视角。
此外,我们发现部分外泌体标志物如TSG101和CD81在exo/IR-MLE中的表达水平升高,而CD63的表达水平没有明显的变化,这表明外泌体中蛋白质的差异性变化包含更多信息。有研究结果显示,微小RNA可以参与T细胞功能的调节[17-18]。作为含有丰富核酸和蛋白质的小囊泡,外泌体具有相同的潜力。因此,未来我们会进一步研究受照射后肺上皮细胞分泌的外泌体中蛋白质和核酸物质的差异变化,这可能为该领域的研究提供新的见解。
综上,我们提出了RILI中存在新抗原的可能性的设想。我们假设照射后的肺上皮细胞分泌的外泌体可以激活T细胞,进而破坏正常肺上皮细胞并加重辐射诱导的不良影响。外泌体可作为一种抗原呈递亚细胞,携带来源细胞经照射后暴露的内部抗原,即机体以前从未识别过的新抗原,并通过其本身携带的、与参与抗原呈递相关的共刺激分子,如MHC Ⅰ、MHC Ⅱ、CD80、CD86等,依赖自身远程递送信息的特性,将抗原传递给初始T细胞。而T细胞通过自身表面的T细胞抗原受体和CD28识别抗原信号后发生活化并出现抗原依赖的增殖现象。
本研究的优势在于证明了外泌体作为抗原呈递亚细胞的作用,揭示了T细胞抗原依赖性活化参与RILI的可能,为今后关于RILI的临床研究和治疗提供了新的思路和实验证据,但仍存在局限性。简单的细胞共培养实验不足以完全模拟机体内部环境;关于抗原呈递的所有结论都来自间接而非直接的证据。我们今后的研究方向将着重于RILI过程中特异性活化T细胞的筛选以及新抗原的鉴定等,同时开展外泌体蛋白质组学分析和测序研究,以期为RILI的研究提供新的实验证据,为今后治疗RILI提供新的实验依据。
综上所述,照射后的MLE-12细胞通过外泌体激活T细胞,这种激活可被外泌体抑制剂所抑制。同时,MLE-12细胞经照射后分泌的外泌体中抗原呈递相关分子高表达,这表明激活机制可能是抗原呈递。本研究为RILI的产生和发展提供了新证据,并为未来的临床研究提供了新的视角。
利益冲突 所有作者声明无利益冲突
作者贡献声明 王志鑫负责实验实施、论文的撰写;王美玉、郭浩鑫负责取材、文献的调研与整理;杜丽负责研究方案可行性的调查分析;王易龙负责实验的设计、论文的修订与审核;杨陟华负责研究思路与方案的设计;朱茂祥负责研究命题的提出、论文的修订与终审
60Co γ射线照射的小鼠肺上皮MLE-12细胞分泌的外泌体对T淋巴细胞的活化作用
Exosome-mediated T cell activation by mouse lung epithelial MLE-12 cells irradiated with 60Co γ ray
-
摘要:
目的 探讨小鼠肺上皮MLE-12细胞(简称MLE-12细胞)受到60Co γ射线照射后分泌的外泌体介导的T细胞活化。 方法 将MLE-12细胞分为对照组和60Co γ射线照射组(2、4、6和8 Gy),采用超速离心法分别从其培养液的上清液中提取外泌体,应用透射电子显微镜和纳米颗粒跟踪分析技术确定外泌体的形态结构和数量特征,采用蛋白质印迹法(WB)检测外泌体中溶酶体相关膜蛋白3(CD63)、四次跨膜蛋白28(CD81)、肿瘤易感基因101蛋白(TSG101)、Ⅰ型内质网膜蛋白(Calnexin)的表达,采用流式细胞术(FCM)检测外泌体表面Ⅰ类主要组织相容性复合体(MHC Ⅰ)、Ⅱ类主要组织相容性复合体(MHC Ⅱ)、免疫调节蛋白B7-1(CD80)和免疫调节蛋白B7-2(CD86)的表达水平。将从小鼠脾脏中分离出来的初始T细胞分别与对照组MLE-12细胞(简称NC MLE-12)分泌的外泌体(简称exo/NC-MLE)、6 Gy γ射线照射组的MLE-12细胞(简称IR MLE-12)分泌的外泌体(简称exo/IR-MLE)共培养,采用FCM检测T细胞亚群CD3+、CD4+和CD8+及其活化指标T细胞特定表面糖蛋白CD28和早期活化抗原1(CD69)的变化;将初始T细胞分别与NC MLE-12、IR MLE-12和外泌体抑制剂GW4869处理组的MLE-12细胞共培养,采用FCM检测T细胞亚群CD3+、CD4+和CD8+及其活化指标CD28和CD69的变化。2组间比较采用两独立样本t检验,多组间比较采用方差分析法,组间两两比较采用Bonferroni调整法。 结果 MLE-12细胞分泌的外泌体显示出典型的一面凹陷的茶托样结构,粒径为30~150 nm;WB结果显示,与MLE-12细胞相比,其外泌体中特异性标志物CD63、CD81和TSG101高表达,而阴性标志物Calnexin低表达。与对照组相比,在6 Gy γ射线照射后不同时间,单个MLE-12细胞分泌的外泌体数量于24、48 h时均增加(t=5.36、6.66,均P<0.05);在不同剂量γ 射线照射后24 h,单个MLE-12细胞分泌的外泌体数量增加的现象具有剂量-效应关系,在照射剂量为 6、8 Gy 时,差异均有统计学意义(t=4.14、5.67,均P<0.05)。与exo/NC-MLE相比,exo/IR-MLE中MHCⅠ、MHC Ⅱ、CD81和TSG101的表达水平均升高。FCM结果显示,与exo/NC-MLE相比,exo/IR-MLE中MHC Ⅰ、MHC Ⅱ、CD80和CD86表达水平均升高(t=4.04~6.47,均P<0.05)。与exo/NC-MLE相比,在与exo/IR-MLE共培养的T细胞中,CD3+、CD4+和CD8+ T细胞均出现增殖现象(t=3.08~5.88,均P<0.05),CD28和CD69表达水平均升高(t=3.02~8.65,均P<0.05);外泌体抑制剂GW4869可以抑制IR MLE-12所诱导的T细胞活化(t=3.64~23.03,均P<0.05)。 结论 60Co γ射线照射后的MLE-12细胞分泌的外泌体可以通过抗原呈递激活T细胞。 Abstract:Objective To evaluate T cell activation driven by exosomes from mouse lung epithelial MLE-12 cells (MLE-12 cells) irradiated with 60Co γ ray. Methods MLE-12 cells were divided into a control group and a 60Co γ irradiation group (2, 4, 6, and 8 Gy), and exosomes were extracted from the supernatant of their culture medium by using ultracentrifugation. Nanoparticle tracking analysis and transmission electron microscope were used to determine the morphological structure and quantity of exosomes. The expression of lysosomal associated membrane protein (CD63), tetraspanin (CD81), tumor susceptibility gene (TSG101), and type Ⅰ endoplasmic reticulum protein (Calnexin) in exosomes were identified by Western blot (WB). Flow cytometry (FCM) was used to detect the expression of major histocompatibility complex class Ⅰ (MHC Ⅰ), major histocompatibility complex class Ⅱ (MHC Ⅱ), immune regulatory protein B7-1 (CD80), and immune regulatory protein B7-2 (CD86) on the surface of exosomes. Naive T cells isolated from mouse spleens were cocultured with exosomes (exo/NC MLE) secreted by MLE-12 cells in the control group (NC MLE-12) and exosomes (exo/IR MLE) secreted by MLE-12 cells in the 6 Gy 60Co γ irradiation group (IR MLE-12), respectively. FCM was used to detect the changes of T cell subsets CD3+, CD4+, and CD8+ and their activated proliferation indicators T cell specific surface glycoprotein CD28 and early activation antigen 1 (CD69). Naive T cells were incubated with NC MLE-12, IR MLE-12, and MLE-12 cells from exosome inhibitor GW4869-treated groups, respectively. FCM was used to detect the changes of T cell subsets CD3+, CD4+, and CD8+ and their activation indicators CD28 and CD69. Independent samples t-test was used for comparison between two groups. Analysis of variance was used to compare multiple groups. Bonferroni adjustment was applied for pairwise comparison between two groups. Results The exosomes produced from MLE-12 cells showed a typical saucer-like structure, with a particle size of 30–150 nm. WB results showed that the exosomes specific markers CD63, CD81, and TSG101 were highly expressed in exosomes, but the negative marker Calnexin was low in expression, compared with the MLE-12 cells. Compared with the control group, at different times after 6 Gy γ ray irradiation, the number of exosomes secreted by a single MLE-12 cell increased at 24 and 48 hours (t=5.36, 6.66, both P<0.05). The phenomenon of an increase in the number of exosomes secreted by a single MLE-12 cell 24 hours after irradiation with different doses of γ rays has a dose-effect relationship, and the difference is statistically significant at doses of 6 and 8 Gy (t=4.14, 5.67, both P<0.05) after the MLE-12 cells were irradiated with γ ray. The expression levels of MHC Ⅰ, MHC Ⅱ, CD81, and TSG101 increased in exo/IR-MLE compared with exo/NC-MLE. FCM results showed that the expression levels of MHC Ⅰ, MHC Ⅱ, CD80, and CD86 increased in exo/IR-MLE compared with exo/NC-MLE (t=4.04–6.47, all P<0.05). Compared with the exo/NC-MLE, in the T cells cocultured with exo/IR-MLE, the CD3+, CD4+, and CD8+ T cells all proliferated (t=3.08–5.88, all P<0.05), and the expression levels of CD28 and CD69 increased (t=3.02–8.65, all P<0.05). The exosome inhibitor GW4869 can suppress T cell activation induced by IR MLE-12 (t=3.64–23.03, all P<0.05). Conclusion Exosomes from MLE-12 cells irradiated with 60Co γ ray could activate T cells through antigen presentation. -
Key words:
- Radiation /
- Alveolar epithelial cells /
- Exosomes /
- T-lymphocytes /
- Antigen presentation
-
图 1 γ射线照射前后小鼠肺上皮MLE-12细胞分泌的外泌体情况
Figure 1. Exosomes secreted by mouse lung epithelial MLE-12 cells before and after irradiated with γ ray
图 2 免疫磁珠分选初始T细胞的纯度和活力
Figure 2. Purity and vitality of naive T cells sorted by immunomagnetic beads
图 3 γ射线照射的小鼠肺上皮MLE-12细胞分泌的外泌体诱导T细胞活化
Figure 3. Activation of T cells by exosomes secreted by mouse lung epithelial MLE-12 cells irradiated with γ ray
图 4 抗原呈递相关分子在γ射线照射前后小鼠肺上皮MLE-12细胞分泌的外泌体中的表达
Figure 4. Expression of antigen presentation related molecules in exosomes secreted by mouse lung epithelial MLE-12 cells before and after irradiated with γ ray
-
[1] Giuranno L, Ient J, De Ruysscher D, et al. Radiation-induced lung injury (RILI)[J/OL]. Front Oncol, 2019, 9: 877[2023-04-02]. https://www.frontiersin.org/articles/10.3389/fonc.2019.00877/full. DOI: 10.3389/fonc.2019.00877. [2] Zhang ZF, Zhou JL, Verma V, et al. Crossed pathways for radiation-induced and immunotherapy-related lung injury[J/OL]. Front Immunol, 2021, 12: 774807[2023-04-02]. https://www.frontiersin.org/articles/10.3389/fimmu.2021.774807/full. DOI: 10.3389/fimmu.2021.774807. [3] Yan YJ, Fu JM, Kowalchuk RO, et al. Exploration of radiation-induced lung injury, from mechanism to treatment: a narrative review[J]. Transl Lung Cancer Res, 2022, 11(2): 307−322. DOI: 10.21037/tlcr-22-108. [4] Guo TT, Zou LQ, Ni JJ, et al. Regulatory T cells: an emerging player in radiation-induced lung injury[J/OL]. Front Immunol, 2020, 11: 1769[2023-04-02]. https://www.frontiersin.org/articles/10.3389/fimmu.2020.01769/full. DOI: 10.3389/fimmu.2020.01769. [5] Cui WC, Zhang P, Hankey KG, et al. AEOL 10150 alleviates radiation-induced innate immune responses in non-human primate lung tissue[J]. Health Phys, 2021, 121(4): 331−344. DOI: 10.1097/hp.0000000000001443. [6] 李倩, 耿爽, 鄢成名, 等. DC细胞在辐射损伤抗原递呈及T细胞活化中的作用[J]. 中国辐射卫生, 2022, 31(6): 657−662, 668. DOI: 10.13491/j.issn.1004-714X.2022.06.003.
Li Q, Geng S, Yan CM, et al. Antigen presentation and T cell activation by dendritic cells in radiation damage[J]. Chin J Radiol Health, 2022, 31(6): 657−662, 668. DOI: 10.13491/j.issn.1004-714X.2022.06.003.[7] Ikhlas S, Usman A, Kim D, et al. Exosomes/microvesicles target SARS-CoV-2 via innate and RNA-induced immunity with PIWI-piRNA system[J/OL]. Life Sci Alliance, 2022, 5(3): e202101240[2023-04-02]. https://www.life-science-alliance.org/content/5/3/e202101240. DOI: 10.26508/lsa.202101240. [8] Liu BW, Jin Y, Yang JY, et al. Extracellular vesicles from lung tissue drive bone marrow neutrophil recruitment in inflammation[J/OL]. J Extracell Vesicles, 2022, 11(5): e12223[2023-04-02]. https://onlinelibrary.wiley.com/doi/10.1002/jev2.12223. DOI: 10.1002/jev2.12223. [9] Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines[J/OL]. J Extracell Vesicles, 2018, 7(1): 1535750[2023-04-02]. https://www.tandfonline.com/doi/full/10.1080/20013078.2018.1535750. DOI: 10.1080/20013078.2018.1535750. [10] Hill GR, Koyama M. Cytokines and costimulation in acute graft-versus-host disease[J]. Blood, 2020, 136(4): 418−428. DOI: 10.1182/blood.2019000952. [11] Reddy M, Eirikis E, Davis C, et al. Comparative analysis of lymphocyte activation marker expression and cytokine secretion profile in stimulated human peripheral blood mononuclear cell cultures: an in vitro model to monitor cellular immune function[J]. J Immunol Methods, 2004, 293(1/2): 127−142. DOI: 10.1016/j.jim.2004.07.006. [12] Chattopadhyay PK, Roederer M. Good cell, bad cell: flow cytometry reveals T-cell subsets important in HIV disease[J]. Cytometry A, 2010, 77A(7): 614−622. DOI: 10.1002/cyto.a.20905. [13] Berg J, Halvorsen AR, Bengtson MB, et al. Circulating T cell activation and exhaustion markers are associated with radiation pneumonitis and poor survival in non-small-cell lung cancer[J/OL]. Front Immunol, 2022, 13: 875152[2023-04-02]. https://www.frontiersin.org/articles/10.3389/fimmu.2022.875152/full. DOI: 10.3389/fimmu.2022.875152. [14] 郗停停, 耿爽, 孙泽文, 等. γ射线胸部照射小鼠早期肺组织的免疫细胞反应[J]. 国际放射医学核医学杂志, 2020, 44(5): 286−290. DOI: 10.3760/cma.j.cn121381-202003038-00025.
Xi TT, Geng S, Sun ZW, et al. Early response of immune-related T cells in the lung tissue of mice exposed to gamma rays in the chest[J]. Int J Radiat Med Nucl Med, 2020, 44(5): 286−290. DOI: 10.3760/cma.j.cn121381-202003038-00025.[15] Kulshreshtha A, Ahmad T, Agrawal A, et al. Proinflammatory role of epithelial cell-derived exosomes in allergic airway inflammation[J]. J Allergy Clin Immunol, 2013, 131(4): 1194−1203. DOI: 10.1016/j.jaci.2012.12.1565. [16] Admyre C, Grunewald J, Thyberg J, et al. Exosomes with major histocompatibility complex class Ⅱ and co-stimulatory molecules are present in human BAL fluid[J]. Eur Respir J, 2003, 22(4): 578−583. DOI: 10.1183/09031936.03.00041703. [17] Shahbaz S, Okoye I, Blevins G, et al. Elevated ATP via enhanced miRNA-30b, 30c, and 30e downregulates the expression of CD73 in CD8+ T cells of HIV-infected individuals[J]. PLoS Pathog, 2022, 18(3): e1010378. DOI: 10.1371/journal.ppat.1010378. [18] Xu YY, Liu ZC, Lv LX, et al. MiRNA-340-5p mediates the functional and infiltrative promotion of tumor-infiltrating CD8+ T lymphocytes in human diffuse large B cell lymphoma[J]. J Exp Clin Cancer Res, 2020, 39(1): 238. DOI: 10.1186/s13046-020-01752-2. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 1669
- HTML全文浏览量: 1256
- PDF下载量: 13
