


 下载:
下载:


-
在全球女性恶性肿瘤中,宫颈癌的发病率和病死率均居第4位[1]。2020年,全球有60.4万宫颈癌新发病例和34.2万宫颈癌死亡病例;中国约有11.0万例宫颈癌患者,其中包括5.9万死亡病例[2]。目前,宫颈癌的治疗方法包括手术、化疗和放疗等[3]。宫颈癌Ⅰ B3~ⅣB期患者均需行放疗,然而约30%接受放疗的患者会出现肿瘤复发[4]。因此,研究影响宫颈癌细胞在放疗后DNA损伤修复的分子机制对提高宫颈癌的放疗效果具有重要意义。
表皮生长因子受体(epidermal growth factor receptor,EGFR)是由原癌基因c-erbB1编码的Ⅰ型跨膜酪氨酸激酶受体(TKR),其由1 210 个氨基酸构成,相对分子质量约为170 000。EGFR分为3个部分:与配体结合的细胞外结构域、跨膜结构域和含有酪氨酸激酶活性的细胞内结构域。EGFR被认为与多种癌症的预后有关,如头颈癌、肺癌、食道癌和胃癌[5]。有研究报道,EGFR信号通路在细胞的生长、增殖和分化等生理过程中发挥重要作用,其过表达或扩增在宫颈癌中发挥调控细胞辐射抗性的作用[6]。
核转录因子E2相关因子2(nuclear factor-E2-related factor 2,NRF2)在保护细胞免受氧化、亲电等多种环境因素威胁,维持暴露在辐射或药物下的细胞内的氧化还原动态平衡方面具有重要作用。有研究结果证明,肿瘤细胞中NRF2的过表达通常与辐射和化学耐药性有关[7]。有文献报道,在黑色素瘤细胞中,NRF2可以促进EGFR的激活,而在口腔癌细胞中EGFR可以促进NRF2的激活[8-9]。但在宫颈癌细胞中,关于EGFR和NRF2在辐射诱导的DNA损伤修复中的关系目前鲜见报道。本研究旨在探讨辐射后的宫颈癌HeLa细胞中EGFR和NRF2对DNA损伤修复的影响及其二者之间的相互关系,为指导宫颈癌的放疗提供理论依据。
-
通过cBioPortal(
http://www.cbioportal.org )对癌症基因组图谱(TCGA)数据库中宫颈癌患者的数据进行分析。 -
甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体、β-Tubulin抗体、Lamin B 抗体、NRF2抗体、辣根过氧化物酶偶联亲和山羊抗小鼠和兔免疫球蛋白G、Cy3标记的山羊抗鼠免疫球蛋白G购于美国Proteintech公司;EGFR抗体购于美国Santa Cruz Biotechnology公司;p-共济失调毛细血管扩张突变基因Rad3相关激酶(ataxia telangiectasia mutated gene and Rad3-related kinase,ATR) (Thr1989)抗体购于美国GeneTex公司;p-细胞周期检查点激酶1(checkpoint kinase 1,CHK1)(Ser345)和血红素氧合酶1(heme oxygenase-1,HO-1)抗体购于美国Cell Signaling Technology公司;磷酸化组蛋白H2A变异体(phosphorylated histone 2A variant,γ-H2AX) (Ser139)抗体购自英国Abcam公司;小干扰RNA(small interfering RNA,siRNA)购于苏州吉玛基因股份有限公司;RNAimax转染试剂、NE-PER 细胞核和细胞质提取试剂购自美国Thermo Fisher公司;胎牛血清(FBS)购于日本HAKATA公司;DMEM液体培养基购于美国HyClone公司;4%组织固定液、牛血清白蛋白(BSA)、Opti MEM培养基购自美国Gibco公司;PBS缓冲液、Loading buffer、Tween-20、碘化丙锭(PI)溶液多聚甲醛、聚乙二醇辛基苯基醚(Triton X-100)和胰蛋白酶-乙二胺四乙酸(EDTA)消化液(0.25%)购自北京索莱宝科技有限公司;快速RNA提取试剂盒购于湖南艾科瑞生物工程有限公司;逆转录试剂盒购自北京宝日医生物技术公司;4′,6-二脒基-2-苯基吲哚(DAPI)封片剂购于北京Vectorlabs公司;三乙醇胺缓冲液(tris buffered saline,TBS)为天津市放射医学与分子核医学重点实验室自制;微型离心管(eppendorf,EP)购于天津本生健康科技公司;低温台式离心机(5424R型)购于德国Eppendorf公司;DMI3000B倒置荧光显微镜购自德国徕卡公司;37℃ 恒温培养箱购自上海力申科学仪器有限公司;CFXConnect实时定量PCR仪购自美国Bio-Rad 公司;Gammacell®40 Exactor 137Cs γ 射线照射源购自加拿大Best Theratronics公司。
-
人宫颈癌HeLa细胞由中国国医学科学院基础医学研究所细胞资源中心提供。人宫颈癌HeLa细胞在含10%胎牛血清的DMEM培养基中,置于5% CO2、37℃的培养箱培养。每隔48 h传代1 次,细胞消化液为胰蛋白酶-乙二胺四乙酸(EDTA)消化液(0.25%)。
细胞经137Cs γ射线照射源进行单次8 Gy照射,吸收剂量率为0.875 Gy/min。
将人宫颈癌HeLa细胞按2种处理方式分组:(1)采用siRNA敲降HeLa细胞中的EGFR,采用137Cs γ射线照射源照射细胞。将HeLa细胞分为对照组(HeLa siCtrl)、敲降EGFR组(HeLa siEGFR)、照射组(HeLa siCtrl+8 Gy)、敲降EGFR+照射组(HeLa siEGFR+8 Gy)。采用免疫荧光实验检测细胞中γ-H2AX foci的数量;采用流式细胞术检测细胞周期;采用实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,RT-qPCR)检测NRF2下游靶基因;采用核质分离实验分离胞质、胞核蛋白;采用蛋白质免疫印迹法检测NRF2、EGFR、HO-1和ATR在Thr1989位点的磷酸化水平,CHK1在Ser345位点的磷酸化水平。(2)采用siRNA敲降HeLa细胞中的NRF2,采用137Cs γ射线照射源照射细胞。将HeLa 细胞分为对照组(HeLa siCtrl)、敲降NRF2组(HeLa siNRF2)、照射组(HeLa siCtrl+8 Gy)、敲降NRF2+照射组(HeLa siNRF2+8 Gy)。采用免疫荧光实验检测细胞中γ-H2AX foci的数量。
-
EGFR的siRNA序列为:5′-GGAGAUAAGUGAUGGAGAUTT-3′
NRF2的siRNA序列为:5′-GGCAUUUCACUAAACACAATT-3′
转染前1 d,将处于对数生长期的HeLa细胞按5×105 个/孔接种于6孔板中,待细胞密度为50%左右时,更换为1 ml新的含10%胎牛血清的DMEM培养基。取4 μl siRNA 和4 μl RNAimax转染试剂分别稀释于200 μl Opti MEM培养基中,将2种溶液混合,使用移液器缓慢吹打混匀,室温放置5 min。将混合液加入含10%胎牛血清的DMEM培养基中,缓慢晃动培养皿使其分布均匀。培养基在37℃、5% CO2的细胞培养箱中孵育8 h,加入1 ml含10%胎牛血清的DMEM培养基(不含抗生素),继续培养至24 h,收集细胞用于后续实验。
-
照射后6 h,收集HeLa细胞至EP管,使用预冷的PBS洗2次,吸干PBS,加入含蛋白酶抑制剂的细胞质提取试剂Ⅰ(CER Ⅰ),涡旋EP管15 s,使细胞颗粒完全悬浮。将EP管放在冰上孵育10 min,向EP管中加入细胞质提取试剂Ⅱ(CER Ⅱ),涡旋EP管5 s;再将EP管在冰上孵育1 min,涡旋5 s。将EP管在离心机(16 000×g,4℃)中离心5 min,立即将上清液(细胞质提取液)转移到另一个干净的EP管中,加入1/3 体积的4×Loading buffer,混匀, 100℃裂解变性10 min。将产生的含核的不溶性(球团)部分悬浮于含有蛋白酶抑制剂的核提取试剂(NER)中,将涡旋调至最高,涡旋15 s。将样本放在冰上孵育,每10 min涡旋15 s,共涡旋5次。将样本在离心机中以最大速度(16 000×g)离心10 s,立即将上清液(核提取液)转移到一个干净的预冷EP管中,加入1/3 体积的4×Loading buffer,于100℃下进行10 min裂解变性。
-
将对数生长期的HeLa细胞按1×105个/孔接种到12孔板中的载玻片上,待细胞完全贴壁后进行8 Gy照射,分别在照射6、12、24 h后取出载玻片,用PBS洗3次,每次5 min;加入4%组织固定液,室温固定20 min,之后再用PBS洗3次,每次5 min;使用预冷的0.3% 聚乙二醇辛基苯基醚(Trinton X-100)室温处理载玻片30 min,PBS洗3次,每次5 min。使用1% BSA(PBS配置)4℃封闭载玻片2 h,在γ-H2AX抗体中4℃孵育12 h,用PBS洗3次,每次5 min。用Cy3标记的山羊抗鼠和兔免疫球蛋白G室温避光孵育载玻片1 h;用PBS洗3次,每次5 min。用含4,6-二脒基-2-苯基吲哚(DAPI)的封片剂封片,将载玻片转移至倒置荧光显微镜下拍照并记录细胞中γ-H2AX foci的数量。
-
采用Western blot法检测EGFR、NRF2、ATR、p-ATR、CHK1、p-CHK1的蛋白表达水平。分别提取各组HeLa细胞的总蛋白,检测蛋白浓度,取30 μg蛋白进行十二烷基硫酸钠-聚丙烯酰胺(SDS-PAGE)电泳分离,转膜,封闭2 h,加入p-ATR(Thr1989)(1∶1 000)、p-CHK1(Ser345)(1∶1 000)、EGFR(1∶1 000)、NRF2(1∶2 000)、GAPDH(1∶5 000)、β-Tubulin(1∶5 000)、Lamin B(1∶5 000)一抗稀释液,4℃ 孵育过夜,使用TBST(tris buffered saline with Tween-20)洗涤30 min,加入二抗(HRP偶联亲和山羊抗小鼠和兔免疫球蛋白G)(1∶5 000),室温孵育2 h,TBST洗涤30 min后显影。
-
采用快速RNA提取试剂盒提取细胞总RNA,用逆转录试剂盒将RNA逆转录为互补DNA(cDNA),检测EGFR、HO-1基因的表达,同时检测GAPDH基因的表达,作为内参。扩增条件:95℃ 激活10 min;95℃ 变性 15 s、60℃ 退火1 min,循环35 次。应用RT-PCR 仪分析荧光强度,采用2−∆∆Ct法对数据进行分析。引物由生工生物工程(上海)股份有限公司合成,序列如下。
GAPDH 引物序列:
正向5′-GTCTCCTCTGACTTCAACAGCG-3′
反向5′-ACCACCCTGTTGCTGTAGCCAA-3′
EGFR引物序列:
正向5′-AGGCACGAGTAACAAGCTCAC-3′
反向5′-ATGAGGACATAACCAGCCACC-3′
HO-1引物序列:
正向 5′-CTTCTTCACCTTCCCCAACA-3′
反向 5′-AGCTCCTGCAACTCCTCAAA-3′
-
将对数生长期的HeLa细胞按5×105 个/孔接种于6孔板中。待细胞完全贴壁后,对细胞进行8 Gy照射,6 h后收集细胞,用PBS清洗2次,去除PBS后用预冷的70%乙醇4℃固定细胞2 h,4℃离心(2 000 r/min,离心半径为10 cm,5 min)去除乙醇,使用预冷的PBS清洗细胞1次,去除PBS,使用碘化丙啶(PI)缓冲液重悬细胞,37℃避光染色30 min,使用流式细胞仪检测细胞周期。
-
应用IBM SPSS Statistics 22.0 软件对数据进行统计学分析。符合正态分布的计量资料以
$ \bar{x} $ ±s 表示,组间比较采用两独立样本t检验(方差齐)。P<0.05 为差异有统计学意义。 -
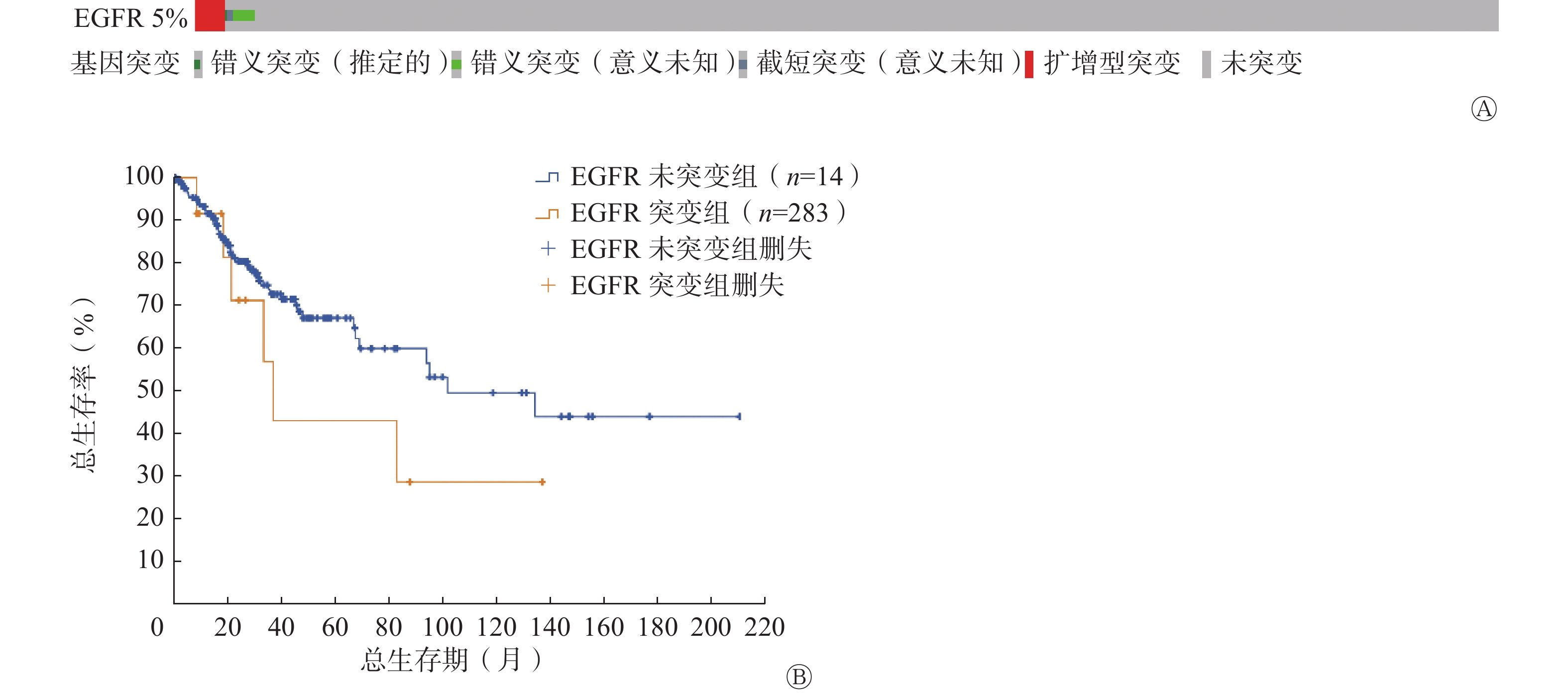
宫颈癌患者中存在5%的EGFR基因突变,其中扩增型突变是主要的突变类型(图1A)。与未发生EGFR基因突变的宫颈癌患者相比,突变患者的生存期更短(图1B)。以上结果提示EGFR对宫颈癌患者的生存率至关重要,可作为宫颈癌的潜在治疗靶标。

图 1 宫颈癌患者的EGFR遗传改变及与生存率的关系
Figure 1. Genetic alterations of epidermal growth factor receptor in cervical cancer patients and its relationship with survival rate
-
细胞免疫荧光实验结果显示,在8 Gy照射后6、12、24 h,HeLa siEGFR组细胞中γ-H2AX foci的数量均多于HeLa siCtrl组[(94.00±1.00)% 对(89.67±2.03)%、(72.33±1.76)% 对(60.00±1.73)%、(43.00±2.31)% 对(26.33±1.20)%],且差异均有统计学意义(均P<0.05)(图2A)。8 Gy照射后6、12、24 h,HeLa siNRF2组细胞中γ-H2AX foci的数量均多于HeLa siCtrl组[(96.67±0.88)%对(89.67±2.03)%、(77.33±1.20)% 对(60.00±1.73)%、(54.33±2.19)% 对(26.33±1.20)%],且差异均有统计学意义(均P<0.05)(图2B)。上述实验结果表明,敲降EGFR或者NRF2均可降低人宫颈癌HeLa细胞的DNA损伤修复能力。

图 2 EGFR、NRF2对人宫颈癌HeLa细胞DNA损伤修复的影响
Figure 2. Effect of epidermal growth factor receptor and nuclear factor-E2-related factor 2 on DNA damage and repair of cervical cancer HeLa cells
-
Western blot实验结果显示,HeLa细胞经8 Gy照射后,EGFR和NRF2蛋白水平均升高,在6 h时达到最大值(图3A),提示在HeLa细胞中EGFR和NRF2可能在调控DNA的损伤修复过程中发挥了重要作用。RT-qPCR实验结果显示,正常条件下敲降EGFR对HO-1表达无明显影响,且差异无统计学意义(t=0.925,P>0.05)。与HeLa siCtrl组细胞比较,HeLa siEGFR+8 Gy的HO-1表达下降66.66%(1.35±0.10对0.45±0.02),且差异有统计学意义(P<0.05),辐射可以导致NRF2靶基因HO-1的表达水平升高(图3B)。流式细胞术结果显示,HeLa siCtrl、HeLa siEGFR、HeLa siCtrl+8 Gy、HeLa siEGFR+8 Gy组细胞的G2周期细胞比例分别是(18.83±0.44)%、(17.07±0.94)%、(44.90±2.81)%、(29.53±2.10)%,与HeLa siCtrl+8 Gy组细胞相比,HeLa siEGFR+8 Gy组的HeLa细胞G2/M期阻滞明显受损,且差异有统计学意义(P<0.05)(图3C)。Western blot实验结果显示,敲降EGFR显著抑制了辐射导致的CHK1的磷酸化和HO-1蛋白水平的升高(图3D)。以上实验结果表明,电离辐射可以引起NRF2和EGFR蛋白水平升高、NRF2下游的DNA损伤响应信号通路和靶蛋白的激活以及G2/M期阻滞。敲降EGFR会抑制电离辐射导致的NRF2下游的ATR-CHK1信号通路的活化和HO-1的激活,并且G2/M期阻滞受损。

图 3 敲降EGFR对HeLa细胞中NRF2下游信号通路及靶蛋白的影响
Figure 3. Effects of epidermal growth factor receptor knockdown on nuclear factor-E2-related factor 2 downstream signaling pathway and target protein in HeLa cells
-
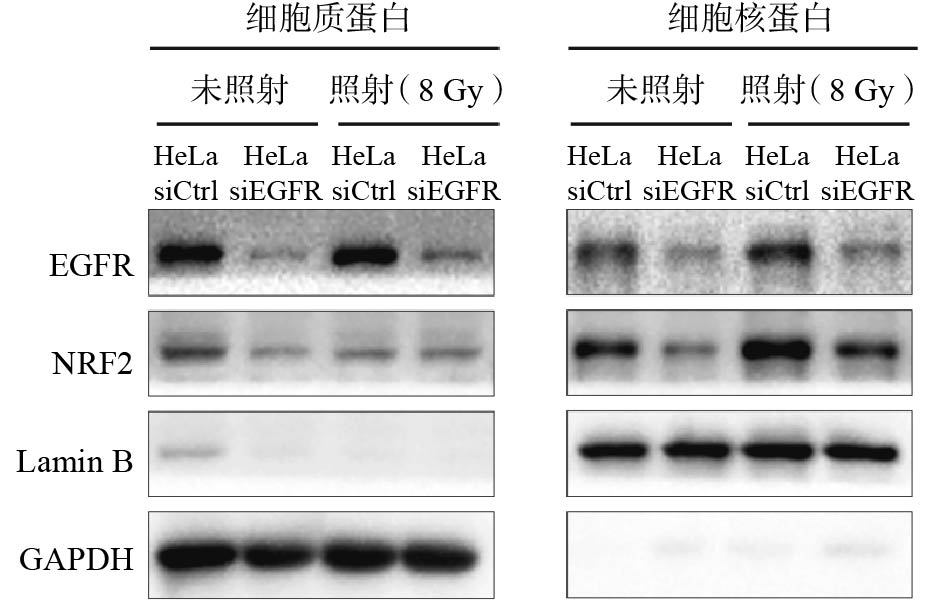
NRF2进入细胞核中才能发挥辐射抵抗的作用,NRF2的核定位对于响应电离辐射至关重要,我们采用核质分离实验来检测EGFR对NRF2核定位的影响,结果如图4所示,敲降EGFR会导致NRF2蛋白水平下降;辐射使在细胞核内定位的NRF2蛋白增加,而敲降EGFR会减少辐射导致的NRF2蛋白在细胞核中的定位,说明EGFR很可能通过促进NRF2的核定位来调控辐射诱导的DNA损伤修复。

图 4 照射和未照射条件下敲降EGFR后HeLa细胞中EGFR、NRF2蛋白在细胞质和细胞核中表达水平的比较
Figure 4. Comparison of epidermal growth factor receptor and nuclear factor-E2-related factor 2 protein expression levels in cytoplasm and nucleus of HeLa cells after knocking down epidermal growth factor receptor under irradiation and non-irradiation conditions
-
放疗是宫颈癌的主要治疗方法之一,一半以上的宫颈癌患者需要接受放疗[10]。近年来, 随着放疗技术的发展,以三维体外照射及三维腔内照射为主的放疗新技术被广泛应用,宫颈癌患者的治疗取得了良好的效果,但辐射抗性仍是导致肿瘤复发或持续的主要因素,因此抑制肿瘤细胞DNA的损伤修复仍非常必要[11]。有研究报道,大约70%的宫颈癌患者中存在EGFR过表达的情况,并且EGFR过表达与疾病特异性生存期(DSS)的缩短相关[12]。本研究对癌症基因组图谱(TCGA)数据库中宫颈癌患者的数据进行分析,发现EGFR突变的宫颈癌患者比例仅为5%,远低于之前的研究结果[12],但疾病特异性生存期(DSS)的缩短确实与EGFR过表达相关,所以EGFR的过表达可能更多发生在mRNA或者蛋白水平。本研究的结果表明,照射后HeLa细胞中EGFR蛋白的表达水平明显升高。为了进一步评估EGFR对宫颈癌细胞DNA损伤修复的影响,我们通过免疫荧光实验观察敲降EGFR后受照HeLa细胞中的DNA损伤情况,结果表明敲降EGFR会降低HeLa细胞的DNA损伤修复能力。
辐射通过直接导致DNA双链断裂或者产生活性氧间接对DNA造成损伤。为了减轻对遗传物质的伤害,细胞会启动DNA损伤反应(DDR)促进对损伤DNA的修复,使细胞周期进程停滞,为修复受损的DNA提供足够的时间[13-14]。通过Western blot、RT-qPCR和流式细胞术实验,我们发现敲降EGFR后,受照射HeLa细胞中ATR-CHK1信号通路的激活受到抑制,HO-1的表达水平降低、细胞G2/M期阻滞受损。这提示EGFR可能通过影响NRF2的功能发挥辐射抵抗作用。
NRF2是基础亮氨酸拉链家族的成员,通过抗氧化反应元件(ARE) 激活细胞内编码抗氧化酶和Ⅱ期解毒酶的基因转录。一般情况下,Kelch样ECH关联蛋白1(Keap1)通过泛素介导的蛋白酶体降解负向调节NRF2活性[15] 。当细胞暴露于氧化、亲电或外来生物刺激时,NRF2会摆脱Kelch样ECH关联蛋白1(Keap1)的控制,转位到细胞核中,与Maf家族的一个小蛋白形成异源二聚体,调控含有抗氧化反应元件(ARE)的下游基因的转录[5, 15]。有研究报道,NRF2的细胞保护功能也可导致对肿瘤细胞的保护,从而促进肿瘤细胞的转化、生长、转移和化疗耐药的形成[16]。有研究结果表明,NRF2通过促进ATR激活和G2/M期阻滞来保持基因组的完整性,DNA发生损伤时,ATR调控的一个主要应答过程是细胞周期阻滞。ATR可通过自身活化并磷酸化蛋白激酶CHK1引起细胞周期阻滞[17]。通过核质分离实验结果,我们发现敲降EGFR明显减少了辐射诱导的细胞核内NRF2蛋白水平的增加。通过免疫荧光实验结果,我们发现敲降NRF2后,HeLa细胞的DNA损伤修复能力降低,表明敲降EGFR会导致NRF2下游信号通路受阻,即电离辐射诱导HeLa细胞DNA损伤修复有赖于EGFR的表达。但目前我们还不确定EGFR是如何调控NRF2在胞内的分布,在今后的研究中,将进一步探索EGFR对NRF2作用的具体机制。
综上所述,本研究采用人宫颈癌HeLa细胞,从细胞及分子水平证实电离辐射后EGFR可促进NRF2蛋白向细胞核内转移,从而激活NRF2下游的ATR/CHK1信号通路及靶基因HO-1的表达,提高细胞的DNA损伤修复能力。因此,我们发现了EGFR在人宫颈癌HeLa细胞辐射抗性中发挥作用的一种可能机制,为靶向EGFR在宫颈癌临床治疗中的应用提供了分子基础及理论依据。
利益冲突 所有作者声明无利益冲突
作者贡献声明 高宇负责实验的实施、论文的撰写与修订;徐畅负责方法的建立、数据的分析、论文的审阅;刘强负责研究命题的设计、论文的修订
EGFR和NRF2对电离辐射导致的DNA损伤修复的影响
The effects of EGFR and NRF2 on DNA damage repair induced by ionizing radiation
-
摘要:
目的 探讨表皮生长因子受体(EGFR)和核转录因子E2相关因子2(NRF2)对人宫颈癌HeLa细胞受到电离辐射损伤后的DNA损伤响应及修复作用。 方法 将人宫颈癌HeLa细胞按2种处理方式分组:(1)采用小干扰RNA敲降HeLa细胞中的EGFR,采用137Cs γ射线照射源照射细胞。将HeLa细胞分为对照组(HeLa siCtrl)、敲降EGFR组(HeLa siEGFR)、照射组(HeLa siCtrl+8 Gy)、敲降EGFR+照射组(HeLa siEGFR+8 Gy)。采用免疫荧光实验(8 Gy照射后6、12、24 h)检测细胞中磷酸化组蛋白H2A变异体(γ-H2AX)foci的数量;采用实时荧光定量聚合酶链式反应(RT-qPCR)检测NRF2下游靶基因;采用流式细胞术检测EGFR对HeLa细胞周期的影响;采用核质分离实验分离HeLa细胞的胞质蛋白和胞核蛋白;采用蛋白质免疫印迹法检测NRF2、EGFR、血红素氧合酶1(HO-1)、共济失调毛细血管扩张突变基因Rad3相关激酶(ATR)Thr1989位点的磷酸化水平、检查点激酶1(CHK1)在Ser345位点的磷酸化水平。(2)采用小干扰RNA敲降HeLa细胞中的NRF2,采用137Cs γ射线照射源照射细胞。将HeLa 细胞分为对照组(HeLa siCtrl)、敲降NRF2组(HeLa siNRF2)、照射组(HeLa siCtrl+8 Gy)、敲降NRF2+照射组(HeLa siNRF2+8 Gy)。采用免疫荧光实验检测细胞中γ-H2AX foci的数量。符合正态分布的计量资料的组间比较采用两独立样本t检验(方差齐)。 结果 (1)8 Gy照射6、12、24 h后,HeLa siEGFR+8 Gy组细胞中γ-H2AX foci的数量均多于HeLa siCtrl组[(94.00±1.00)% 对(89.67±2.03)%、(72.33±1.76)% 对(60.00±1.73)%、(43.00±2.31)% 对(26.33±1.20)%],且差异有统计学意义(t=3.919、4.919、6.402,均P<0.05)。与HeLa siCtrl组比较,HeLa siEGFR+8 Gy组的细胞G2/M期阻滞显著受损[(46.53±3.06)%对(37.90±4.61)%],且差异有统计学意义(t=4.384,P<0.05)。与HeLa siCtrl组比较,HeLa siEGFR+8 Gy组的HO-1表达下降66.66%(1.35±0.10对0.45±0.02),且差异有统计学意义(t=8.782,P<0.05)。敲降EGFR后细胞核内的NRF2蛋白水平降低,辐射引起的NRF2下游ATR-CHK1信号通路活化水平及HO-1蛋白水平均降低。(2)8 Gy照射6、12、24 h后,与HeLa siCtrl组相比,HeLa siNRF2+8 Gy组细胞中γ-H2AX foci的数量均多于HeLa siCtrl组[(96.67±0.88)%对(89.67±2.03)%、(77.33±1.20)% 对(60.00±1.73)%、(54.33±2.19)% 对(26.33±1.20)%],且差异均有统计学意义(t=3.166、4.919、11.220,均P<0.05)。 结论 电离辐射条件下,敲降EGFR可以减少NRF2蛋白入核,抑制ATR-CHK1信号通路激活及下游基因HO-1的表达,降低人宫颈癌HeLa细胞的DNA损伤修复能力。 -
关键词:
- 宫颈肿瘤 /
- DNA损伤 /
- DNA修复 /
- 表皮生长因子受体 /
- 核转录因子E2相关因子2
Abstract:Objective To investigate the effect of epidermal growth factor receptor (EGFR) and nuclear factor-E2-related factor 2(NRF2) on DNA damage response and repair in human cervical cancer HeLa cells after exposure to ionizing radiation. Methods Human cervical cancer HeLa cells were treated in two groups. (1) EGFR was knocked down in HeLa cells using a small interfering RNA, and the cells were irradiated using a 137Cs γ-ray irradiation source. HeLa cells were divided into control group (HeLa siCtrl), knockdown EGFR group (HeLa siEGFR), irradiation group (HeLa siCtrl+8 Gy), and knockdown EGFR+irradiation group (HeLa siEGFR+8 Gy). The number of phosphorylated histone 2A variant (γ-H2AX) foci in the cells was detected by immunofluorescence assay (6, 12, and 24 h after 8 Gy irradiation); the NRF2 downstream target genes were detected by real-time fluorescence quantitative polymerase chain reaction (RT-qPCR) assay; the expression levels of flow cytometry were used to detect the effect of EGFR on HeLa cell cycle; nucleoplasmic separation assay was used to isolate cytoplasmic and cytosolic proteins from HeLa cells; protein immunoblotting was used to detect the phosphorylation levels of NRF2, EGFR, HO-1, ataxia-telangiectasia mutated gene and Rad3-related kinase (ATR) Thr1989 locus, cell cycle checkpoint kinase 1 (p-CHK1), and Ser345 site phosphorylation level. (2) A small interfering RNA was used to knock down NRF2 in HeLa cells, and the cells were irradiated with a 137Cs γ-ray irradiation source. HeLa cells were divided into control group (HeLa siCtrl), knockdown NRF2 group (HeLa siNRF2), irradiation group (HeLa siCtrl+8 Gy), and knockdown NRF2+ irradiation group (HeLa siNRF2+8 Gy). An immunofluorescence assay was used to detect the number of γH2AX foci in HeLa cells. Inter-group comparisons of measures conforming to normal distribution were performed by two independent sample t-tests (chi-squared). Results (1) After 8 Gy irradiation for 6, 12, and 24 h, the number of γH2AX foci in HeLa siEGFR+8 Gy were more than that HeLa siCtrl ((94.00±1.00)% vs. (89.67±2.03)%, (72.33±1.76)% vs. (60.00±1.73)%, (43.00±2.31)% vs. (26.33±1.20)%), and the differences were statistically significant (t=3.919, 4.989, 6.402; all P<0.05). The HeLa siEGFR+8 Gy impaired radiation-induced G2/M phase cell cycle block compared with the HeLa siCtrl, ((46.53±3.06)% vs. (37.90±4.61)%), and the difference was statistically significant (t=4.384, P<0.05). Compared with the HeLa siCtrl, HeLa siEGFR+8 Gy inhibited the radiation induced decrease in HO-1 expression by 66.66%(1.35±0.10 vs. 0.45±0.02), and the difference was statistically significant (t=8.782, P<0.05). The level of NRF2 protein in the nucleus was reduced after knocking down EGFR, and the radiation-induced activation level of the ATR-CHK1 signaling pathway downstream of NRF2 and the level of HO-1 protein were reduced. (2) After 8 Gy irradiation for 6, 12, and 24 h, the number of γH2AX foci in HeLa siNRF2+8 Gy was more than that HeLa siCtrl ((96.67±0.88)% vs. (89.67±2.03)%, (77.33±1.20)% vs. (60.00±1.73)%, (54.33±2.19)% vs. (26.33±1.20)%), and all differences were statistically significant (t=3.166, 4.989, 11.220; all P<0.05). Conclusions Under ionizing radiation conditions, knocking down EGFR can reduce the nuclear translocation of NRF2 protein, inhibit the activation of the ATR-CHK1 signaling pathway and the downstream expression of the HO-1 gene, and decrease the DNA damage repair capacity of human cervical cancer HeLa cells. -
图 1 宫颈癌患者的EGFR遗传改变及与生存率的关系
Figure 1. Genetic alterations of epidermal growth factor receptor in cervical cancer patients and its relationship with survival rate
图 2 EGFR、NRF2对人宫颈癌HeLa细胞DNA损伤修复的影响
Figure 2. Effect of epidermal growth factor receptor and nuclear factor-E2-related factor 2 on DNA damage and repair of cervical cancer HeLa cells
图 3 敲降EGFR对HeLa细胞中NRF2下游信号通路及靶蛋白的影响
Figure 3. Effects of epidermal growth factor receptor knockdown on nuclear factor-E2-related factor 2 downstream signaling pathway and target protein in HeLa cells
图 4 照射和未照射条件下敲降EGFR后HeLa细胞中EGFR、NRF2蛋白在细胞质和细胞核中表达水平的比较
Figure 4. Comparison of epidermal growth factor receptor and nuclear factor-E2-related factor 2 protein expression levels in cytoplasm and nucleus of HeLa cells after knocking down epidermal growth factor receptor under irradiation and non-irradiation conditions
-
[1] Siegel RL, Miller KD, Fuchs HE, et al. Cancer statistics, 2022[J]. CA Cancer J Clin, 2022, 72(1): 7−33. DOI: 10.3322/caac.21708. [2] Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209−249. DOI: 10.3322/caac.21660. [3] Dueñas-González A, Cetina L, Coronel J, et al. The safety of drug treatments for cervical cancer[J]. Expert Opin Drug Saf, 2016, 15(2): 169−180. DOI: 10.1517/14740338.2016.1130127. [4] Tewari KS, Monk BJ. Evidence-based treatment paradigms for management of invasive cervical carcinoma[J]. J Clin Oncol, 2019, 37(27): 2472−2489. DOI: 10.1200/JCO.18.02303. [5] Baselga J. Why the epidermal growth factor receptor? The rationale for cancer therapy[J]. Oncologist, 2002, 7(Suppl 4): S2−8. DOI: 10.1634/theoncologist.7-suppl_4-2. [6] Gaffney DK, Haslam D, Tsodikov A, et al. Epidermal growth factor receptor (EGFR) and vascular endothelial growth factor (VEGF) negatively affect overall survival in carcinoma of the cervix treated with radiotherapy[J]. Int J Radiat Oncol Biol Phys, 2003, 56(4): 922−928. DOI: 10.1016/S0360-3016(03)00209-8. [7] Singh A, Bodas M, Wakabayashi N, et al. Gain of Nrf2 function in non-small-cell lung cancer cells confers radioresistance[J]. Antioxid Redox Signal, 2010, 13(11): 1627−1637. DOI: 10.1089/ars.2010.3219. [8] Kreß JKC, Jessen C, Marquardt A, et al. NRF2 enables EGFR signaling in melanoma cells[J/OL]. Int J Mol Sci, 2021, 22(8): 3803[2023-03-08]. https://www.mdpi.com/1422-0067/22/8/3803. DOI: 10.3390/IJMS22083803. [9] Carrillo-Beltrán D, Muñoz JP, Guerrero-Vásquez N, et al. Human papillomavirus 16 E7 promotes EGFR/PI3K/AKT1/NRF2 signaling pathway contributing to PIR/NF-κB activation in oral cancer cells[J/OL]. Cancers (Basel), 2020, 12(7): 1904[2023-03-08]. https://www.mdpi.com/2072-6694/12/7/1904. DOI: 10.3390/cancers12071904. [10] Cao LJ, Wen H, Feng Z, et al. Distinctive clinicopathologic characteristics and prognosis for different histologic subtypes of early cervical cancer[J]. Int J Gynecol Cancer, 2019, 29(8): 1244−1251. DOI: 10.1136/ijgc-2019-000556. [11] Krause M, Dubrovska A, Linge A, et al. Cancer stem cells: radioresistance, prediction of radiotherapy outcome and specific targets for combined treatments[J]. Adv Drug Deliv Rev, 2017, 109: 63−73. DOI: 10.1016/j.addr.2016.02.002. [12] Schrevel M, Gorter A, Kolkman-Uljee SM, et al. Molecular mechanisms of epidermal growth factor receptor overexpression in patients with cervical cancer[J]. Mod Pathol, 2011, 24(5): 720−728. DOI: 10.1038/modpathol.2010.239. [13] Jackson SP, Bartek J. The DNA-damage response in human biology and disease[J]. Nature, 2009, 461(7267): 1071−1078. DOI: 10.1038/nature08467. [14] Ciccia A, Elledge SJ. The DNA damage response: making it safe to play with knives[J]. Mol Cell, 2010, 40(2): 179−204. DOI: 10.1016/j.molcel.2010.09.019. [15] Chung HC, Ros W, Delord JP, et al. Efficacy and safety of pembrolizumab in previously treated advanced cervical cancer: results from the phase Ⅱ KEYNOTE-158 study[J]. J Clin Oncol, 2019, 37(17): 1470−1478. DOI: 10.1200/JCO.18.01265. [16] Wang YY, Wang Y, Zhang ZB, et al. Mechanism of progestin resistance in endometrial precancer/cancer through Nrf2-AKR1C1 pathway[J/OL]. Oncotarget, 2016, 7(9): 10363−10372[2023-03-08]. https://www.oncotarget.com/article/7004/text. DOI: 10.18632/oncotarget.7004. [17] Sun XH, Wang Y, Ji KH, et al. NRF2 preserves genomic integrity by facilitating ATR activation and G2 cell cycle arrest[J]. Nucleic Acids Res, 2020, 48(16): 9109−9123. DOI: 10.1093/nar/gkaa631. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1722
- HTML全文浏览量: 1218
- PDF下载量: 32
