


 下载:
下载:




-
肺癌是癌症中导致死亡的首要原因[1-2]。约有85%的肺癌患者被诊断为非小细胞肺癌(non-small cell lung cancer,NSCLC),而90%的NSCLC患者病死与肿瘤转移有关[3]。多数NSCLC患者发病时已经发生了多处转移,而未经治疗的转移性患者的中位生存时间仅为4~5个月,1年生存率仅为10%[4]。尽管目前对肺癌转移方面的研究有了新的进展,但肺癌转移患者的预后仍然不乐观。因此,有必要进一步拓宽对NSCLC转移机制的认识,以改善对该病的治疗效果。
放疗被认为是无法进行手术的早期NSCLC患者的主要治疗手段。立体定向放射疗法(stereotactic body radiation therapy,SBRT)可以安全、有效地对无法进行手术的早期NSCLC患者(包括基础肺功能较差的患者)进行治疗[5]。尽管SBRT对早期NSCLC患者的局部控制率很好(3年期为98%、5年期为87%)[6-7],但患者在随访中经常发生远处转移;对于不能手术治疗的Ⅲ期NSCLC患者,化疗联合放疗的试验结果显示,肿瘤控制率或患者生存期并未得到明显改善[8]。因此,目前仍需要探索影响NSCLC辐射敏感性的因素,从而改善NSCLC的放疗效果,特别是在疾病的早期阶段。
富含半胱氨酸型运动神经元蛋白1(cysteine-rich motor neuron 1,CRIM1)是骨形态发生蛋白抑制剂家族中富含半胱氨酸区域的Ⅰ型跨膜糖蛋白,相对分子质量约为130 000,含有1036个氨基酸,由胞质区、跨膜域和胞外域组成。在CRIM1蛋白的羧基末端,有一个跨膜结构域和一个相对较短的胞质区,胞外域包括位于氨基末端的信号肽序列、胰岛素样生长因子结合蛋白样结构域和6个高度保守的富含半胱氨酸的重复序列,胞外域可以从细胞中释放,形成分泌型CRIM1[9-10]。在本研究中,我们通过体内外实验探讨CRIM1对NSCLC辐射敏感性和转移的影响,并初步探索其作用机制。
-
胎牛血清购自上海传秋生物科技有限公司,1640培养液购自美国HyClone公司,慢病毒颗粒购自上海吉凯公司,小干扰RNA(small interfering RNA,siRNA)购自苏州吉玛基因股份有限公司,Trizol试剂购自美国赛默飞世尔科技公司,磷酸化组蛋白H2AX(γ-H2AX)抗体购自美国Abcam公司,Cy3标记的山羊抗鼠免疫球蛋白G购自美国Proteintech公司,Matrigel基质胶和Transwell小室购自美国康宁公司,正常熔点琼脂糖购自美国Biowest公司,低熔点琼脂糖购自美国Promega公司,Super GelRed染液购自苏州宇恒生物科技公司,逆转录试剂盒购自北京宝日医生物技术公司,结晶紫、多聚甲醛、聚乙二醇辛基苯基醚(Triton X-100)和纤连蛋白购自北京索莱宝科技有限公司,细胞计数试剂盒8(CCK-8)购自大连美仑生物技术有限公司。DMI3000B倒置荧光显微镜购自德国徕卡公司,37℃恒温培养箱购自上海力申科学仪器有限公司,Gammacell®40 Exactor 137Cs γ射线照射源购自加拿大Best Theratronics公司,CFX Connect实时定量PCR仪购自美国Bio-Rad公司。
-
人NSCLC H460、H358细胞均购自美国模式培养物集存库(ATCC)细胞库,均采用含10%胎牛血清的1640培养液,在含5%CO2的37℃恒温培养箱中培养,待细胞融合度达到80%时进行传代培养。选择处于对数生长期的细胞进行后续实验。
-
慢病毒LV-CRIM1-RNAi(LV:慢病毒载体;RNAi:RNA干扰)和阴性对照LV-CON均由上海吉凯生物公司构建。将H460、H358细胞按照1×105个/孔接种于12孔板中,培养24 h,用慢病毒颗粒(感染复数=10)进行感染。感染12 h后,将含病毒的培养液换为完全培养基。感染96 h后,用嘌呤霉素筛选,后扩大培养。将H460、H358细胞分别分为3组:H460细胞、H460-shCRIM1细胞、H460-shNC细胞和H358细胞、H358-shCRIM1细胞、H358-shNC细胞(其中shCRIM1表示用shRNA敲降CRIM1的表达,shNC表示阴性对照)。
将H358细胞按照1×106个/孔接种于6孔板中,待细胞融合度达到70%时,进行转染。将siRNA转染H358细胞分为2组:H358-NC细胞(转染siRNA-NC,即阴性对照)和H358-siCRIM1细胞(转染siRNA-CRIM1,即CRIMI敲降序列:正义链为5′-CCUUAUUGCUGGCUGCAAUTT-3′;反义链为 5′-AUUGCAGCCAGCAAUAAGGTT-3′)。转染24 h后弃去原有培养液,加入新鲜完全培养基,继续培养24 h后用Trizol试剂裂解细胞,采用荧光定量PCR法检测干扰效率。
-
雄性BALB/c-nu裸鼠30只,6~8周龄,体重(20±2)g,由北京华阜康生物科技股份有限公司提供[许可证号:SCXK(京)2020-0004],饲养于中国医学科学院北京协和医学院放射医学研究所实验动物中心。所有动物在无特定病原体条件下,于恒温(23℃±2℃)、恒湿(45%~50%)、无菌净化屏障系统内饲养,随意采食全价鼠饲料及清洁水。动物实验开展前获得了中国医学科学院北京协和医学院放射医学研究所动物伦理委员会的批准(批准号:IRM-DWLL-2020140)。
将30只裸鼠分为3组,每组10只,分别接种H460、H460-shCRIM1、H460-shNC细胞。将处于对数生长期的上述3种细胞经胰酶消化,离心(115×g)5 min后弃上清,用PBS洗涤2次并离心(115×g)5 min,用Matrigel基质胶混合制备成5×106个/mL的细胞悬液。于BALB/c-nu裸鼠右腋前线第二肋上缘,右肺第二叶注射100 μL细胞悬液,2周后将小鼠全部处死并取肺脏。
-
使用137Cs γ射线照射源对细胞进行照射,剂量率为1 Gy/min。
-
采用Trizol法提取细胞总RNA,用逆转录试剂盒将RNA逆转录为cDNA,检测相关基因的表达,包括CRIM1、连接黏附分子2(junctional adhesion molecule 2,JAM2)、连接蛋白3(nectin cell adhesion molecule 3,NECTIN3)和紧密连接蛋白4(claudin 4,CLDN4)等基因,同时检测甘油醛-3-磷酸脱氢酶(GAPDH)基因的表达,并作为内参。扩增条件:95℃激活10 min;95℃变性 15 s、60℃退火1 min,循环35次。应用实时定量PCR仪分析荧光强度,数据采用 2−ΔΔCt 法进行分析。引物由上海生工生物工程有限公司合成,序列如下。
GAPDH引物序列:
正向5′-GTCTCCTCTGACTTCAACAGCG-3′;
反向5′-ACCACCCTGTTGCTGTAGCCAA-3′;
CRIM1引物序列:
正向5′-ACGCGATCACAATGGTTGTCGG-3′;
反向5′-GGCATCAGTAAGGAAACCGAAGG-3′;
NECTIN3引物序列:
正向5′- ATTCCCGCTTGGAAATGCCCAG-3′;
反向5′- GCTGCTACTGTTTCATTTCCTCC-3′;
CLDN4引物序列:
正向5′- AGTGCAAGGTGTACGACTCGCT-3′;
反向5′- CGCTTTCATCCTCCAGGCAGTT-3′;
JAM2引物序列:
正向5′- AGACTTGGCTCCCAAAGCACCA-3′;
反向5′- TTCCCAGGACACCTGCGATATC-3′。
-
分别将500个H460、H460-shCRIM1、H460-shNC细胞接种至6孔培养板中,待细胞贴壁后,分别进行0、1、2、4 Gy照射,照射时间分别为0、1、2、4 min,并继续培养至出现肉眼可见的克隆,共计7 d。弃掉培养基,用结晶紫染液染色。肉眼进行克隆计数,以每团细胞数>50个作为1个克隆。根据公式进行计算,细胞克隆形成率=照射细胞克隆数/未照射(0 Gy照射)同种细胞克隆数×100%。
-
将H460、H460-shNC、H460-shCRIM1细胞接种至载玻片上24 h后,行6、8、12 Gy照射,照射时间分别为6、8、12 min,12 h后取出载玻片,PBS清洗,4%多聚甲醛室温下固定15 min,0.3% Triton X-100室温下处理细胞15 min,5%牛血清蛋白室温封闭2 h,加入磷酸化组蛋白H2AX(γ-H2AX)一抗30 μL,4℃孵育过夜,PBS清洗3次,加入二抗(Cy3标记的山羊抗鼠免疫球蛋白G)。室温孵育1 h,PBS清洗,4′,6-二脒基-2-苯基吲哚(DAPI)染色后于荧光显微镜下拍照并记录细胞中磷酸化组蛋白H2AX(γ-H2AX)foci数。
-
收集H460、H460-shNC、H460-shCRIM1细胞,用PBS冲洗2次,并用PBS重悬,调节细胞悬液密度至4×105个/mL,经8 Gy照射后立即铺板。彗星实验板(自制)每孔铺入0.75%正常熔点琼脂糖凝胶约500 μL,待凝结后,将30 μL细胞悬液与70 μL 0.75%低熔点琼脂糖凝胶混匀并滴于上层铺平,将彗星实验板完全浸没在细胞裂解液中,于4℃裂解2.5 h。再将彗星实验板移入装有Tris-硼酸缓冲液的电泳槽中进行DNA解旋20 min(30 V电压下电泳20 min),电泳结束后用中和缓冲液中和20 min。PBS冲洗彗星实验板后用Super GelRed染液染色,在荧光显微镜下拍照。应用CASP 6.0软件(
http://www.casp-program.org )分析图像;casp_1.2.3b1彗星分析软件(波兰弗罗茨瓦夫大学)分析olive尾距。 -
用胰蛋白酶消化H460、H460-shCRIM1、H460-shNC细胞和H358、H358-shCRIM1、H358-shNC细胞,再用无血清培养基稀释。在Transwell小室的上室中加入5×104个细胞,在下室中加入600 μL含10%胎牛血清的1640培养基,将上室放入24孔板后放入细胞培养箱继续培养。24 h后取出上室,用乙醇配制的0.5%结晶紫固定染色30 min,PBS清洗,擦去上室内残留细胞,晾干后用倒置显微镜拍照。
-
在实验前12 h,将纤连蛋白涂布于96孔板中;将胰蛋白酶消化后的H460、H460-shCRIM1、H460-shNC细胞和H358、H358-shCRIM1、H358-shNC细胞铺至上述96孔板中,37℃孵育20 min,用无血清1640培养基洗去未黏附细胞,再加入无血清1640培养基,37℃孵育4 h后加入细胞计数试剂盒8(CCK-8)试剂,检测450 nm波长下的吸光度(A450值),根据吸光度值判定细胞黏附能力。
-
将小鼠处死后迅速取出肺脏,经生理盐水漂洗后,用4%中性甲醛溶液固定,在左右肺的最大截面处进行组织病理切片,然后用苏木精-伊红染色,在显微镜下观察肺组织中肿瘤的分布情况。
-
用Trizol法提取H460-shCRIM1、H460-shNC细胞总RNA,每种细胞设3个重复样品,并进行测序(委托广州基迪奥公司完成)。对得到的数据进行相关分析,以差异表达倍数>2、校准后P<0.05为筛选标准,筛选得到差异基因。利用基因本体论(gene ontology,GO)数据库对全部差异基因进行注释,京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)数据库对全部差异基因进行通路分析,注释与通路分析均使用北京大学KOBAS系统(
http://kobas.cbi.pku.edu.cn )进行,采用超几何分布计算差异基因显著富集的GO 条目和KEGG通路,P<0.05为显著富集。筛选得到目的基因后进行荧光定量PCR检测,具体方法同1.6节。 -
应用IBM SPSS Statistics 22.0软件进行统计学分析。符合正态分布的计量资料以
$\bar x $ ±s表示,在方差齐的条件下,组间比较采用两独立样本t检验。P<0.05为差异有统计学意义。 -
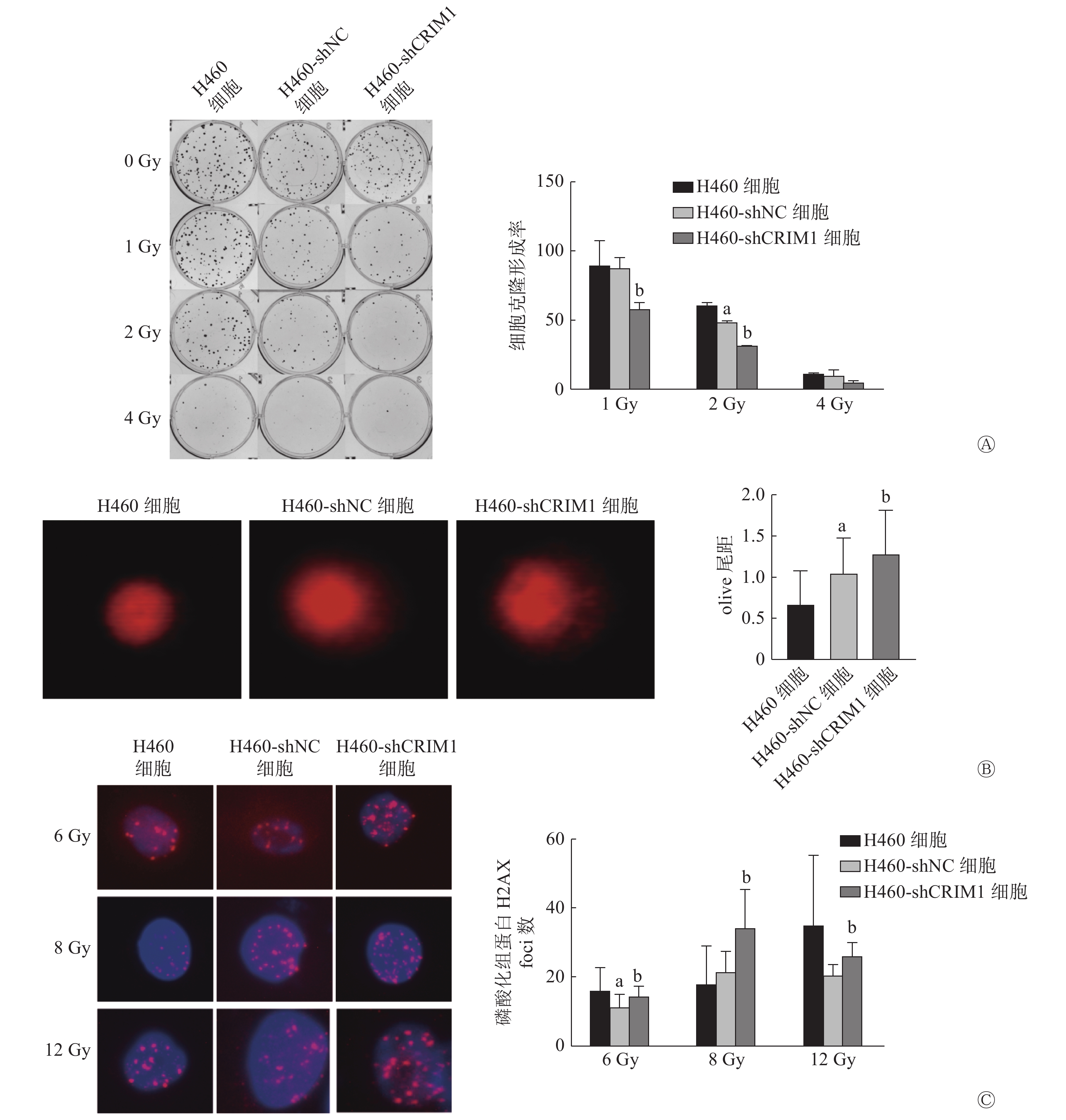
细胞克隆形成实验结果显示,H460-shNC细胞和H460-shCRIM1细胞在接受1、2、4 Gy照射后的克隆形成率分别为(87.04±8.04)%、(48.23±1.22)%、(9.45±4.21)%和(58.01±4.39)%、(31.43±0.08)%、(4.79±1.54)%。与H460-shNC细胞相比,1、2 Gy照射后H460-shCRIM1细胞克隆形成率明显降低,且差异均有统计学意义(t=4.48、19.50,均P<0.05),4 Gy照射后H460-shCRIM1细胞克隆形成率也呈现出下降的趋势(图1A)。彗星实验结果显示,H460-shCRIM1细胞的olive尾距长于H460-shNC细胞,且差异有统计学意义(1.27±0.54对1.05±0.42,t=2.14,P<0.05)(图1B)。细胞免疫荧光实验结果显示,H460-shCRIM1细胞受照后的磷酸化组蛋白H2AX(γ-H2AX)foci数多于H460-shNC细胞(6 Gy:14.33±2.81对11.00±3.92;8 Gy:34.00±11.14对21.17±6.15;12 Gy:25.80±3.96对20.17±3.31),且差异均有统计学意义(t=2.45、5.52、2.47,均P<0.05)(图1C)。由此可见,敲降H460细胞内CRIM1的表达可以增强其辐射敏感性。H358细胞由于克隆形成能力较差,未检测其辐射敏感性。
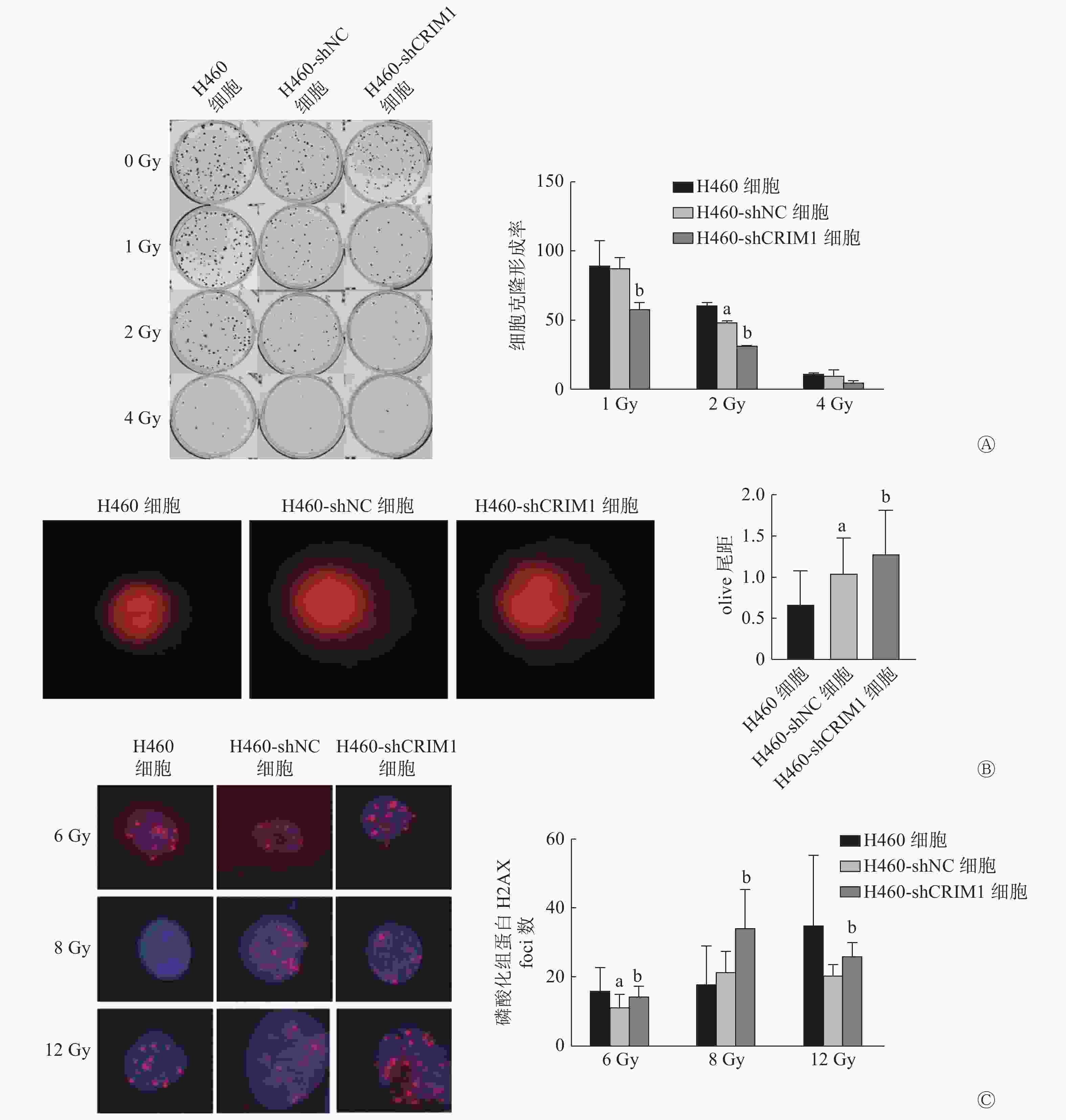
图 1 CRIM1对非小细胞肺癌H460细胞辐射敏感性的影响
Figure 1. Effect of cysteine-rich moter neuron 1 on radiosensitivity of non-small cell lung cancer H460 cell
-
由图2可见,shRNA可以从转录水平敲降H460、H358细胞中CRIM1 mRNA的相对表达量(t=35.35、22.50,均P<0.05)。Transwell实验结果显示,H460-shCRIM1细胞的迁移率较H460-shNC细胞明显升高(t=4.73,P<0.05),黏附能力下降(t=2.86,P<0.05)(图3A);H358-shCRIM1细胞的迁移率较H358-shNC细胞明显升高(t=10.19,P<0.05),黏附能力下降(t=3.66,P<0.05)(图3B)。小鼠肺内原位接种肿瘤实验结果显示,接种后2周,H460-shCRIM1细胞的肺内转移灶明显多于H460-shNC细胞(图3C、D),且在纵隔和胸膜下出现大量的转移灶。
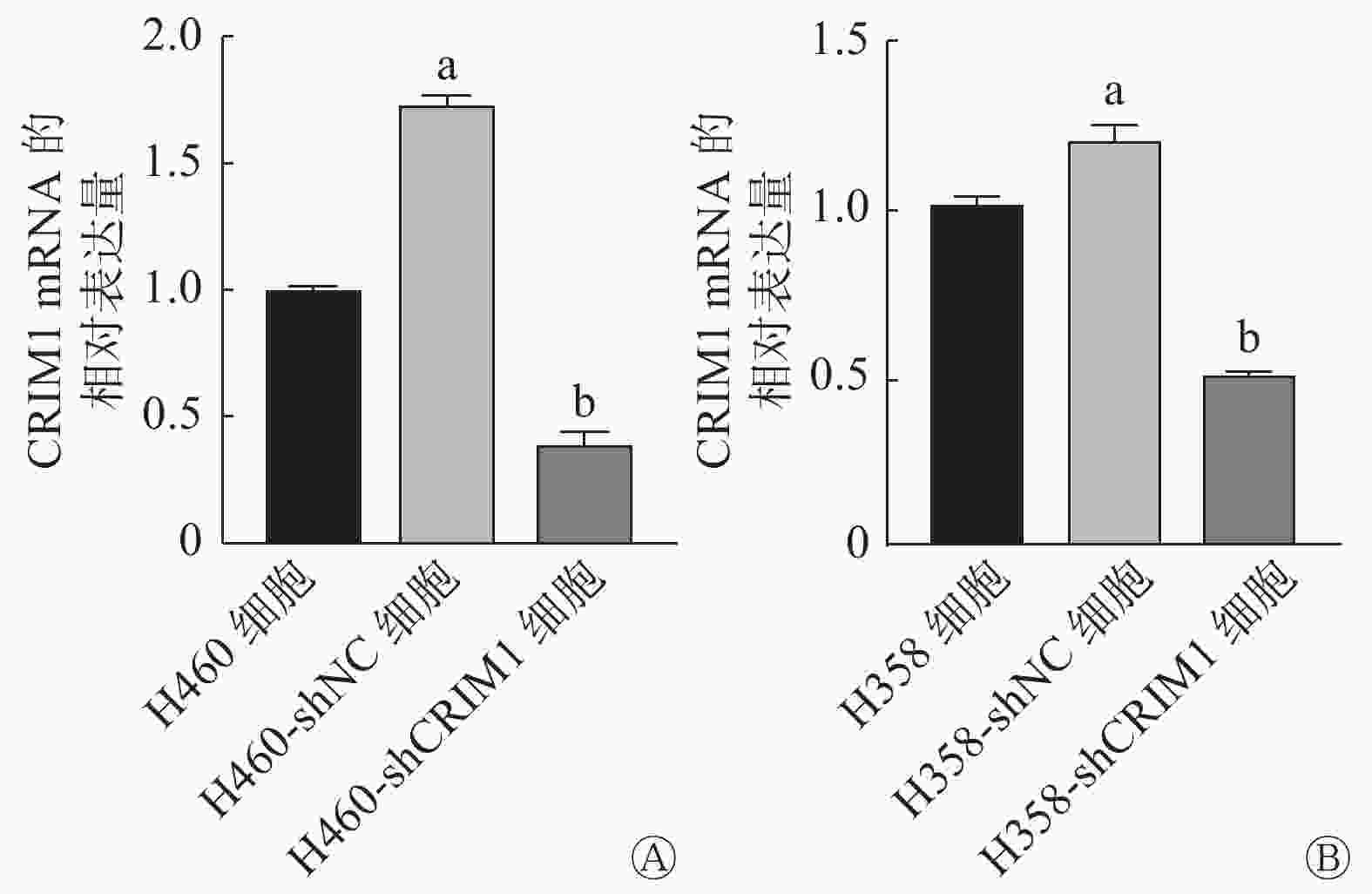
图 2 短发夹RNA敲降后非小细胞肺癌H460、H358细胞中CRIMI mRNA的相对表达量
Figure 2. Effect of short hairpin RNA on cysteine-rich motor neuron 1 expression in non-small cell lung cancer H460 and H358 cells
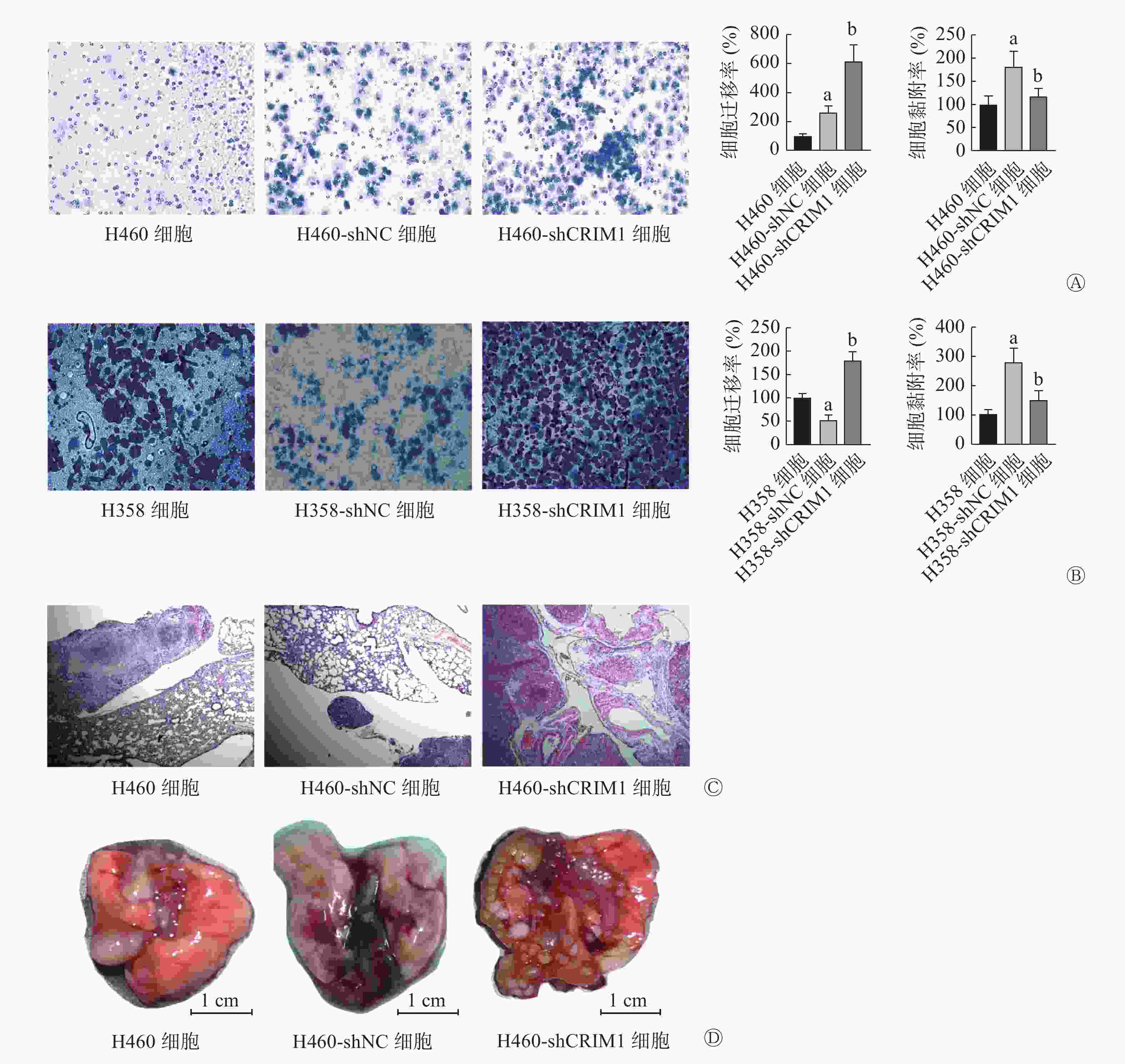
图 3 CRIM1对非小细胞肺癌H460、H358细胞转移的影响
Figure 3. Effect of cysteine-rich motor neuron 1 on the metastasis of non-small cell lung cancer H460 and H358 cells
-
转录组测序结果显示,通过主成分分析(PCA)结果发现,降维分析后H460-shCRIM1细胞和H460-shNC细胞的样本聚集度较高(图4A)。与H460-shNC细胞相比,H460-shCRIM1细胞中表达上调基因58个、下调基因39个。对所有97个基因和39个下调基因的KEGG富集分析结果(图4B)显示,其显著变化的通路均富集在细胞黏附分子通路上,富集的基因是参与细胞黏附连接的NECTIN3、CLDN4基因和构成细胞紧密连接的JAM2基因,其相对表达量在H460-shCRIM1细胞中均有所下降。
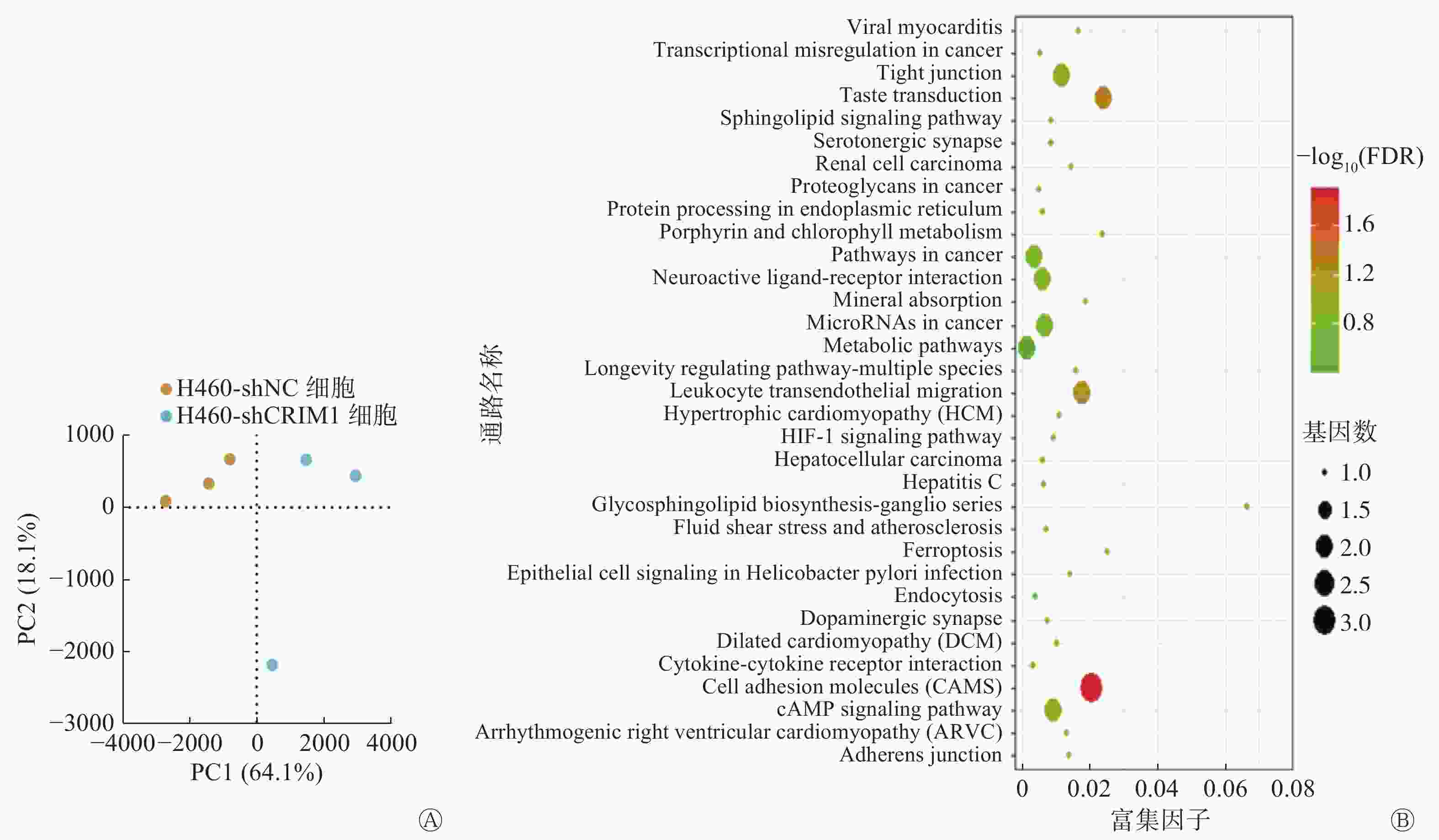
图 4 敲降非小细胞肺癌H460细胞中CRIM1的表达后的转录组基因分析
Figure 4. Transcriptome gene analysis after reducing cysteine-rich motor neuron 1 expression in non-small cell lung cancer H460 cell
-
由图5可见,与H460-shNC细胞比较,H460-shCRIM1细胞中的JAM2、NECTIN3和CLDN4基因的相对表达量分别下降了86.66%、49.35%、30.27%(t=47.52、7.47、18.98,均P<0.05)(图5A)。与H358-siNC细胞比较,H358-siCRIM1细胞中的JAM2、NECTIN3和CLDN4基因的相对表达量分别下降了36.60%、31.70%、50.00%(t=7.40、7.10、16.56,均P<0.05)(图5C)。由此可见,CRIM1的表达降低可能会导致NSCLC中细胞黏附分子转录水平的降低。
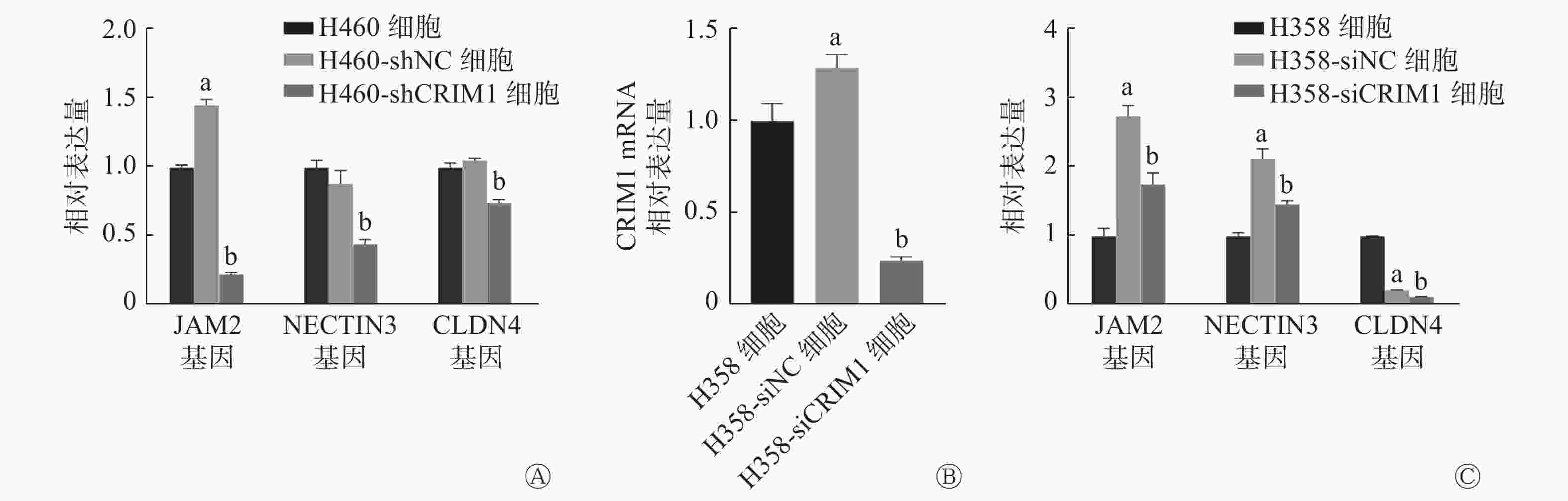
图 5 敲降非小细胞肺癌H460、H358细胞中CRIM1的表达对细胞黏附相关基因表达的影响
Figure 5. Expression of cell adhesion-related genes after reducing cysteine-rich motor neuron 1 expression in non-small cell lung cancer H460 and H358 cells
-
放疗尽管在NSCLC的治疗上取得了一定进展,但进一步提高NSCLC的辐射敏感性仍然是非常有必要的。Hurov等[11]对人骨肉瘤U2OS细胞进行全基因组RNA干扰分析,筛选结果显示CRIM1可能与降低U2OS细胞的辐射敏感性相关。本研究结果显示,敲降CRIM1的H460-shCRIM1细胞照射后的克隆形成能力降低。DNA是辐射的直接作用靶点,因此DNA的损伤程度可间接反映细胞对辐射的敏感性。为了进一步评估CRIM1对NSCLC H460细胞辐射敏感性的影响,我们通过慧星实验和细胞免疫荧光实验观察了敲降CRIM1后受照H460细胞的DNA损伤情况,结果也表明敲降CRIM1会增加照射对H460细胞DNA的损伤程度,增强其辐射敏感性。随后,我们分析了转录组学结果,但未显示出与辐射敏感性紧密相关的信号通路的改变。推测CRIM1可能通过不依赖于转录水平变化的方式来影响H460细胞的辐射敏感性。
转移是肿瘤治疗失败的主要原因,而肿瘤的迁移、侵袭、黏附和血管形成等能力将直接影响肿瘤的转移。本研究主要从迁移和黏附2个角度阐述了CRIM1对NSCLC细胞转移能力的影响。首先采用shRNA慢病毒感染与siRNA瞬时转染2种NSCLC H460和H358细胞,分别采用Transwell实验和细胞黏附实验评估了细胞的迁移和黏附能力,结果表明敲降CRIM1可以促进NSCLC细胞的迁移和黏附,进而促进NSCLC细胞的转移;且肺内原位接种H460细胞的体内实验结果也表明,CRIM1低表达的肺癌细胞更易形成多发的转移灶。由此证实CRIM1具有抑制NSCLC细胞转移的作用。CRIM1与肿瘤转移的关系的研究已有报道。Ogasawara等[12]的研究结果表明,CRIM1可抑制肾癌细胞的侵袭和转移;Zeng等[13]的研究结果显示,敲降CRIM1可抑制肺癌A549细胞在体外的迁移。本研究结果与Ogasawara等[12]的结果一致,但与Zeng等[13]的研究结果相反。选用细胞系的不同可能是造成这一差异的原因之一。另外,我们还比较了NSCLC细胞和多种小细胞肺癌细胞中CRIM1的表达,结果发现CRIM1在小细胞肺癌细胞中普遍低表达(研究结果尚未发表)。而这与小细胞肺癌易转移、对辐射敏感的特性相吻合,这也与本研究的结果相吻合。目前,我们正在就CRIM1对小细胞肺癌转移的影响进行探讨,以期深入了解CRIM1与肺癌转移的关系。
关于CRIM1影响NSCLC细胞转移的具体机制,我们分析了敲降CRIM1和阴性对照NSCLC细胞的转录组学的变化。结果发现,细胞黏附分子通路在H460-shNC细胞和H460-shCRIM1细胞之间的差异有统计学意义,通路中的3个细胞黏附分子JAM2、CLDN4、NECTIN3在敲降CRIM1的H460和H358细胞中的相对表达量均明显下调。这提示CRIM1可能是通过影响NSCLC细胞间的黏附发挥其改变肺癌转移的作用。对非洲爪蟾的神经板发育的研究结果也表明,CRIM1主要通过与钙黏着蛋白和β-catenin形成复合物来影响细胞连接复合物的形成和稳定,而该功能对于细胞黏附至关重要[14]。细胞黏附是由桥粒、黏附连接和紧密连接等组成的细胞间锚定和信号传递的场所,用于维持和形成正常的细胞间黏附。细胞黏附的失调在恶性肿瘤的转化和转移过程中起着至关重要的作用[15-16]。细胞黏附连接除发挥锚定作用外,对肌动蛋白细胞骨架、细胞内信号和转录过程的调节均有重要意义。因此,连接通讯系统的失调在肿瘤侵袭中具有特殊意义[17-18]。紧密连接及其相关黏附因子的失调也与肿瘤的转化和侵袭有很大关系。多项研究结果显示,在乳腺癌、肝癌、子宫内膜癌和卵巢癌中,CLDN失调明显会导致肿瘤的发展和转移的潜能增加[19-20]。
因此,我们后期将使用电镜观察肿瘤组织中的细胞间连接,包括黏附连接和紧密连接的变化,以进一步确定CRIM1对细胞连接的影响。对于CRIM1是如何影响细胞黏附分子的水平及其具体的机制,还有待进一步的研究。
综上所述,抑制CRIM1能够增强NSCLC H460细胞的辐射敏感性;CRIM1可能通过影响肿瘤细胞连接的方式影响NSCLC的转移。
利益冲突 本研究由署名作者按以下贡献声明独立开展,不涉及任何利益冲突。
作者贡献声明 李科君负责实验的实施、论文的撰写与修订;赵雯月负责细胞学的实验;李娜负责动物的实验、数据的获取;王彦负责方法的建立、数据的分析;刘强负责论文的审阅;杜利清负责研究命题的设计、论文的修订。
CRIM1对非小细胞肺癌辐射敏感性和转移的影响
Effects of CRIM1 on radiosensitivity and metastasis of non-small cell lung cancer
-
摘要:
目的 探讨富含半胱氨酸型运动神经元蛋白1(CRIM1)对非小细胞肺癌(NSCLC)辐射敏感性和转移的影响及可能的机制。 方法 采用短发夹RNA(shRNA)敲降NSCLC H460、H358细胞中CRIM1的表达,并将H460、H358细胞分别分为3组:H460细胞、H460-shCRIM1细胞、H460-shNC细胞和H358细胞、H358-shCRIM1细胞、H358-shNC细胞,其中shCRIM1表示用shRNA敲降CRIM1的表达,shNC表示阴性对照。采用小干扰RNA(siRNA)敲降H358细胞中CRIM1的表达,并将细胞分为2组:H358-siNC细胞和H358-siCRIM1细胞。采用细胞克隆形成实验(照射剂量为0、1、2、4 Gy)、慧星实验(照射剂量为8 Gy)和细胞免疫荧光实验(照射剂量为6、8、12 Gy)观察CRIM1对H460细胞辐射敏感性的影响。采用Transwell实验、细胞黏附实验观察CRIM1对H460、H358细胞转移的影响。构建小鼠H460原位肿瘤模型,采用组织病理学检查评估裸鼠肺内原位接种肿瘤转移情况。采用转录组学分析法探讨CRIM1影响NSCLC细胞转移的可能机制。组间比较采用两独立样本t检验。 结果 克隆形成实验结果显示,与H460-shNC细胞相比,1、2 Gy照射后H460-shCRIM1细胞的克隆形成率明显降低 [1 Gy:(87.04±8.04)%对(58.01±4.39)%;2 Gy:(48.23±1.22)%对(31.43±0.08)%],且差异均有统计学意义(t=4.48、19.50,均P<0.05)。慧星实验结果显示,8 Gy照射后H460-shCRIM1细胞的olive尾距长于H460-shNC细胞,且差异有统计学意义(1.27±0.54对1.05±0.42,t=2.14,P<0.05)。细胞免疫荧光实验结果显示,H460-shCRIM1细胞受照后磷酸化组蛋白H2AX foci数多于H460-shNC细胞(6 Gy:14.33±2.81对11.00±3.92;8 Gy:34.00±11.14对21.17±6.15;12 Gy:25.80±3.96对20.17±3.31),且差异均有统计学意义(t=2.45、5.52、2.47,均P<0.05)。Transwell实验结果显示,H460-shCRIM1、H358-shCRIM1细胞的迁移率分别较H460-shNC、H358-shNC细胞明显升高,且差异均有统计学意义(t=4.73、10.19,均P<0.05)。细胞黏附实验结果显示,H460-shCRIM1、H358-shCRIM1细胞的黏附能力分别较H460-shNC、H358-shNC细胞下降,且差异均有统计学意义(t=2.86、3.66,均P<0.05)。组织病理学检查结果显示,原位接种H460-shCRIM1细胞的裸鼠肺内转移灶明显多于H460-shNC细胞。转录组学分析结果显示,筛选出的细胞黏附相关基因连接黏附分子2(JAM2)、连接蛋白3(NECTIN3)和紧密连接蛋白4(CLDN4)在H460-shCRIM1、H358-siCRIM1细胞中的表达均下降;并在体外实验中得到验证,其中,H460-shCRIM1细胞相较于H460-shNC细胞分别下降了86.66%、49.35%、30.27%(t=47.52、7.47、18.98,均P<0.05),H358-siCRIM1细胞相较于H358-siNC细胞分别下降了36.60%、31.70%、50.00%(t=7.40、7.10、16.56,均P<0.05)。 结论 抑制CRIM1可增强NSCLC细胞H460的辐射敏感性,CRIM1可能通过影响肿瘤细胞连接的方式影响NSCLC的转移。 -
关键词:
- 癌,非小细胞肺 /
- 辐射耐受性 /
- 肿瘤转移 /
- 富含半胱氨酸型运动神经元蛋白1
Abstract:Objective To investigate the effects of cysteine-rich motor neuron protein 1 (CRIM1) on radiosensitivity and metastasis of non-small cell lung cancer (NSCLC) and its possible mechanisms. Methods Short hairpin RNA (shRNA) was used to knock down the expression level of CRIM1 in NSCLC cells H460 and H358, and H460 and H358 cells were divided into three groups respectively: H460, H460-shCRIM1, H460-shNC, and H358, H358-shCRIM1, H358-shNC, where shCRIM1 indicates the knockdown of CRIM1 expression level by shRNA and shNC indicates the negative control. Small interfering RNA was used to knock down the expression of CRIM1 in H358 cells, and the cells were divided into two groups: H358-siNC and H358-siCRIM1. Clone formation assay (irradiation dose of 0, 1, 2, and 4 Gy), comet assay (irradiation dose of 8 Gy), and cellular immunofluorescence assay (irradiation dose of 6, 8, and 12 Gy) were used in observing the effect of CRIM1 on the radiosensitivity of H460 cells. Transwell and cell adhesion assays were used in observing the effect of CRIM1 on cell metastasis in vitro. A mouse in situ tumor model was constructed, and the metastasis of tumors inoculated in situ into nude mouse lungs was assessed using histopathological examination, and transcriptomic analysis was used in exploring the possible effects of CRIM1 on the mechanism of NSCLC cell metastasis. Independent samples t-test was used in comparing groups. Results Clone formation assay showed statistically significant differences between the clone formation rates of H460-shNC and H460-shCRIM1 after 1 and 2 Gy of irradiation ((1 Gy: (87.04±8.04)% vs. (58.01±4.39)%, t=4.48, P<0.05; 2 Gy: (48.23±1.22)% vs. (31.43±0.08)%, t=19.50, P<0.05)). The results of comet assay showed that the olive tail moment of H460-shCRIM1 cells after 8 Gy of irradiation was longer than that of H460-shNC cells, and the difference was statistically significant (1.27±0.54 vs. 1.05±0.42, t=2.14, P<0.05). The results of cell immunofluorescence experiments showed that the number of phosphorylated histone H2AX foci was higher in H460-shCRIM1 cells than in H460-shNC after irradiation (6 Gy: 14.33±2.81 vs. 11.00±3.92; 8 Gy: 34.00±11.14 vs. 21.17±6.15; 12 Gy: 25.80±3.96 vs. 20.17±3.31), and the difference was statistically significant (t=2.45, 5.52, 2.47; all P<0.05). Transwell assay showed that the mobility rates of H460-shCRIM1 and H358-shCRIM1 were significantly higher than those of H460-shNC and H358-shNC, respectively, and the differences were statistically significant (t=4.73, 10.19; both P<0.05). The results of cell adhesion assay showed that the adhesion ability of H460-shCRIM1 and H358-shCRIM1 decreased relative to that of H460-shNC and H358-shNC, respectively, and the difference was statistically significant (t=2.86, 3.66; both P<0.05). Histopathological examination results showed the in situ inoculation of H460-shCRIM1 into nude mice. Intrapulmonary metastases of H460-shCRIM1 were more extensive than those in H460-shNC. Transcriptomic analysis showed that the expression levels of the screened cell adhesion-related genes junctional adhesion molecule 2 (JAM2), nectin cell adhesion molecule 3 (NECTIN3), and claudin 4 (CLDN4) decreased in H460-shCRIM1 and H358-siCRIM1, and this result was verified in in vitro experiments. The expression levels in H460-shCRIM1 decreased by 86.66% (JAM2), 49.35% (NECTIN3), and 30.27% (CLDN4) compared with those in H460-shNC (t=47.52, 7.47, 18.98; all P<0.05), and in H358-siCRIM1 decreased by 36.60% (JAM2), 31.70% (NECTIN3), and 50.00% (CLDN4) compared with those in H358-siNC (t=7.40, 7.10, 16.56; all P<0.05). Conclusion The inhibition of CRIM1 enhances the radiosensitivity of NSCLC cells H460, and CRIM1 may influence NSCLC metastasis by affecting tumor cell junctions. -
图 1 CRIM1对非小细胞肺癌H460细胞辐射敏感性的影响
Figure 1. Effect of cysteine-rich moter neuron 1 on radiosensitivity of non-small cell lung cancer H460 cell
图 2 短发夹RNA敲降后非小细胞肺癌H460、H358细胞中CRIMI mRNA的相对表达量
Figure 2. Effect of short hairpin RNA on cysteine-rich motor neuron 1 expression in non-small cell lung cancer H460 and H358 cells
图 3 CRIM1对非小细胞肺癌H460、H358细胞转移的影响
Figure 3. Effect of cysteine-rich motor neuron 1 on the metastasis of non-small cell lung cancer H460 and H358 cells
图 4 敲降非小细胞肺癌H460细胞中CRIM1的表达后的转录组基因分析
Figure 4. Transcriptome gene analysis after reducing cysteine-rich motor neuron 1 expression in non-small cell lung cancer H460 cell
-
[1] McIntyre A, Ganti AK. Lung cancer—A global perspective[J]. J Surg Oncol, 2017, 115(5): 550−554. DOI: 10.1002/jso.24532. [2] Cao MM, Chen WQ. Epidemiology of lung cancer in China[J]. Thorac Cancer, 2019, 10(1): 3−7. DOI: 10.1111/1759-7714.12916. [3] Herbst RS, Morgensztern D, Boshoff C. The biology and management of non-small cell lung cancer[J]. Nature, 2018, 553(7689): 446−454. DOI: 10.1038/nature25183. [4] Rapp E, Pater JL, Willan A, et al. Chemotherapy can prolong survival in patients with advanced non-small-cell lung cancer—report of a Canadian multicenter randomized trial[J]. J Clin Oncol, 1988, 6(4): 633−641. DOI: 10.1200/jco.1988.6.4.633. [5] Stanic S, Paulus R, Timmerman RD, et al. No clinically significant changes in pulmonary function following stereotactic body radiation therapy for early-stage peripheral non-small cell lung cancer: an analysis of RTOG 0236[J]. Int J Radiat Oncol Biol Phys, 2014, 88(5): 1092−1099. DOI: 10.1016/j.ijrobp.2013.12.050. [6] Onishi H, Shirato H, Nagata Y, et al. Stereotactic body radiotherapy (SBRT) for operable stage I non-small-cell lung cancer: can SBRT be comparable to surgery?[J]. Int J Radiat Oncol Biol Phys, 2011, 81(5): 1352−1358. DOI: 10.1016/j.ijrobp.2009.07.1751. [7] Miyasaka Y, Komatsu S, Abe T, et al. Comparison of oncologic outcomes between carbon ion radiotherapy and stereotactic body radiotherapy for early-stage non-small cell lung cancer[J/OL]. Cancers, 2021, 13(2): 176[2021-01-25]. https://www.mdpi.com/2072-6694/13/2/176. DOI: 10.3390/cancers13020176. [8] Ahn JS, Ahn YC, Kim JH, et al. Multinational randomized phase Ⅲ trial with or without consolidation chemotherapy using docetaxel and cisplatin after concurrent chemoradiation in inoperable stage Ⅲ non-small-cell lung cancer: KCSG-LU05-04[J]. J Clin Oncol, 2015, 33(24): 2660−2666. DOI: 10.1200/jco.2014.60.0130. [9] Wilkinson L, Kolle G, Wen DY, et al. CRIM1 regulates the rate of processing and delivery of bone morphogenetic proteins to the cell surface[J]. J Biol Chem, 2003, 278(36): 34181−34188. DOI: 10.1074/jbc.M301247200. [10] Furuichi T, Tsukamoto M, Saito M, et al. Crim1C140S mutant mice reveal the importance of cysteine 140 in the internal region 1 of CRIM1 for its physiological functions[J]. Mamm Genome, 2019, 30(11/12): 329−338. DOI: 10.1007/s00335-019-09822-3. [11] Hurov KE, Cotta-Ramusino C, Elledge SJ. A genetic screen identifies the triple T complex required for DNA damage signaling and ATM and ATR stability[J]. Genes Dev, 2010, 24(17): 1939−1950. DOI: 10.1101/gad.1934210. [12] Ogasawara N, Kudo T, Sato M, et al. Reduction of membrane protein CRIM1 decreases E-cadherin and increases claudin-1 and MMPs, enhancing the migration and invasion of renal carcinoma cells[J]. Biol Pharm Bull, 2018, 41(4): 604−611. DOI: 10.1248/bpb.b17-00990. [13] Zeng H, Zhang Y, Yi Q, et al. CRIM1, a newfound cancer-related player, regulates the adhesion and migration of lung cancer cells[J]. Growth Factors, 2015, 33(5/6): 384−392. DOI: 10.3109/08977194.2015.1119132. [14] Ponferrada VG, Fan JQ, Vallance JE, et al. CRIM1 complexes with β-catenin and cadherins, stabilizes cell-cell junctions and is critical for neural morphogenesis[J/OL]. PLoS One, 2012, 7(3): e32635[2021-01-25]. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0032635. DOI: 10.1371/journal.pone.0032635. [15] Feng ZR, Yu QX, Zhang T, et al. Updates on mechanistic insights and targeting of tumour metastasis[J]. J Cell Mol Med, 2020, 24(3): 2076−2086. DOI: 10.1111/jcmm.14931. [16] Bhat AA, Uppada S, Achkar IW, et al. Tight junction proteins and signaling pathways in cancer and inflammation: a functional crosstalk[J/OL]. Front Physiol, 2019, 9: 1942[2021-01-25]. https://www.frontiersin.org/articles/10.3389/fphys.2018.01942/full. DOI: 10.3389/fphys.2018.01942. [17] Rübsam M, Broussard JA, Wickström SA, et al. Adherens junctions and desmosomes coordinate mechanics and signaling to orchestrate tissue morphogenesis and function: an evolutionary perspective[J/OL]. Cold Spring Harb Perspect Biol, 2018, 10(11): a029207[2021-01-25]. https://cshperspectives.cshlp.org/content/10/11/a029207. DOI: 10.1101/cshperspect.a029207. [18] Citi S. The mechanobiology of tight junctions[J]. Biophys Rev, 2019, 11(5): 783−793. DOI: 10.1007/s12551-019-00582-7. [19] Youssef MY, Mohamed MA. Could e-cadherin and CD10 expression be used to differentiate between atypical endometrial hyperplasia and endometrial carcinoma?[J]. Int J Gynecol Pathol, 2019, 38(2): 128−137. DOI: 10.1097/pgp.0000000000000492. [20] Zeisel MB, Dhawan P, Baumert TF. Tight junction proteins in gastrointestinal and liver disease[J]. Gut, 2019, 68(3): 547−561. DOI: 10.1136/gutjnl-2018-316906. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5200
- HTML全文浏览量: 3831
- PDF下载量: 29
