


 下载:
下载:


-
宫颈癌是在全球女性疾病中普遍存在的、发病率和致死率最高的癌症之一[1]。放疗适用于各期宫颈癌的治疗,是局部晚期宫颈癌不可替代的治疗手段,而辐射抗性是导致宫颈癌放疗后局部复发和远处转移的重要原因。
缺氧导致的肿瘤辐射抗性是影响放疗疗效的重要因素,缺氧诱导因子α家族(hypoxia-inducible factor-α family,HIFs)在其中发挥着关键作用。缺氧作为应激源可以调节去泛素化酶(deubiquitinase,DUB),泛素羧基末端水解酶L5(ubiquitin carboxyl-terminal hydrolase isozyme L5,UCHL5)是DUB泛素羧基末端水解酶家族成员,它可影响肿瘤细胞的DNA损伤修复[2]、周期调控[3]、凋亡[4]和增殖等生物进程。本研究旨在观察缺氧对宫颈癌Hela细胞UCHL5表达的影响,进一步探讨UCHL5对宫颈癌Hela细胞辐射敏感性的作用。
-
人宫颈癌Hela细胞和人肾上皮293T细胞购自中国科学院上海细胞生物研究所。
-
高糖DMEM培养基、胰蛋白酶、二甲基亚砜(DMSO)、P8340蛋白酶抑制剂Cocktail和牛血清白蛋白均购自美国Sigma公司;胎牛血清购自杭州四季青生物工程材料有限公司;嘌呤霉素(Puromycin)购自美国Gibco公司;MTT购自上海索莱宝生物科技有限公司;11668-019 lipofectamine 2000、31985-062 Opti-MEM和10296010 TRIzol™均购自美国Invitrogen公司;Dual-Luciferase® Reporter Assay System E1910报告基因检测系统购自美国Promega公司。过表达UCHL5组、过表达对照组、沉默UCHL5组、沉默对照组慢病毒载体和聚凝胺(polybrene)均由上海吉凯基因化学技术有限公司构建;mRNA提取试剂盒和Taqman mRNA试剂盒均购自天根生化科技(北京)有限公司;G4507 姬姆萨染液购自德国Merck公司;559763 PE Annexin V凋亡检测试剂盒购自美国BD公司;RIPA细胞裂解液购自中国(北京)Solarbio公司;BCA蛋白检测试剂盒购自中国碧云天公司;增强的化学发光(ECL)试剂和聚偏氟乙烯膜(PVDF)均购自美国Millipore公司;抗UCHL5抗体、抗缺氧诱导因子1α(hypoxia inducible factor-1α,HIF-1α)抗体均购自美国Abcam公司;抗β-actin抗体、抗Bcl-2抗体和抗兔HRP标记的二抗均购自美国Cell Signaling Technology公司;其他试剂均为国产分析纯。60Co辐照源由空军军医大学辐照中心提供,辐照剂量率为1 Gy/min;CytoFLEX流式细胞分析仪购自美国Beckman Coulter公司。
-
常氧培养:将Hela细胞置于高糖DMEM培养基中(含10% 胎牛血清),在37℃、5% CO2 培养箱中培养。乏氧培养:将Hela细胞置于37℃、1% O2、5% CO2 、94% N2中,分别培养0、1、2、4、6、8、24 h后提取细胞总蛋白。
-
慢病毒载体委托上海吉凯基因化学技术有限公司构建。转录本分为空白对照组、过表达对照组、过表达 UCHL5 组转录本 1~4 和沉默对照组、沉默 UCHL5组转录本 1~2。将处于对数生长期的Hela细胞用0.25%的胰蛋白酶消化后计数,将密度为4×104个/mL的细胞接种于6孔培养板中,37℃培养过夜。培养至细胞融合度为70%~80%时,根据病毒感染复数值计算病毒和聚凝胺浓度,然后将其加入至细胞培养基中,感染12 h后,更换新鲜培养液,继续培养48 h后,更换含2 μg/mL嘌呤霉素的DMEM完全培养基,筛选细胞1周。
-
使用TRIzol™和mRNA提取试剂盒提取细胞总RNA,实时定量PCR检测UCHL5 mRNA的表达。引物由杨凌天润奥科生物科技有限公司合成,具体如下。UCHL5正向:GAGTGGTGCCTCATGGAAAG,反向:CAAGTCGAGTCCTGAACC;β-actin正向:CCTGGGCATGGAGTCCTGTG,反向:TCTTCATTGTGCTGGGTGCC。PCR反应条件:50℃、2 min,95℃、10 min,95℃、15 s,60℃、1 min,共40个循环;使用循环阈值量化对照组和实验组细胞的UCHL5 mRNA水平。采用2−△△CT方法比较对照组和实验组的差异,β肌动蛋白(β-actin)作为内参基因。
-
实验分组为过表达对照组、过表达UCHL5组、沉默对照组和沉默UCHL5组。取处于对数生长期的Hela细胞,分别以100、100、200、400个/孔接种于6孔培养板中,24 h后分别接受单次0、2、4、6 Gy γ射线照射,培养2周后弃去培养基,加甲醇固定15 min,姬姆萨染色20 min,计数50个细胞以上的克隆并计算克隆形成率。克隆形成率=(克隆形成数/铺板细胞数)×100%。
-
取处于对数生长期的过表达和沉默UCHL5细胞及其各自对照组细胞,分别以1000个/孔的数量接种于96孔培养板中,设置5个复孔;分别给予0、2、4、6、8、10 Gy γ射线照射,于0、3、5、7 d后每孔加入20 μL 5 g/L的MTT,37℃培养4 h后吸去培养基,加入150 μL 二甲基亚砜,置于摇床上室温避光震荡10 min,使其充分溶解,酶标仪检测490 nm处的吸光度(optical density,OD)值。细胞增殖率=(实验组OD值−空白对照组OD值)/(对照组OD值−空白对照组OD值)×100%。
-
分别取处于对数生长期的过表达UCHL5组、过表达对照组、沉默UCHL5组和沉默对照组细胞,以5×105个/孔接种于6孔培养板;分别给予0 Gy和8 Gy γ射线照射48 h后,以1000 r/min离心10 min(离心半径为7 cm)后,收集样本,用PBS洗2次,加入559763 PE Annexin V 凋亡检测试剂盒里的Binding Buffer和Annexin V-PE 混匀后,避光反应20 min,检测前5 min再加入7-氨基放线菌素D(7-AAD),用流式细胞仪检测细胞凋亡情况。
-
JASPAR靶基因预测数据库(
http://jaspar.genereg.net )检测结果发现,HIF-1α在UCHL5的启动子区存在互补结合位点。为进一步验证HIF-1α和UCHL5之间的靶向关系,构建了野生型(pGL4.10-UCHL5 Promoter Wt)和突变型(pGL4.10-UCHL5 Promoter Mut)UCHL5荧光素酶重组质粒,将野生型UCHL5(UCHL5Wt)质粒和突变型UCHL5(UCHL5Mut)质粒分别与过表达HIF-1α对照组及过表达HIF-1α组的质粒共转染人肾上皮293T细胞48 h后,采用Dual-Luciferase®报告基因检测系统检测各处理组293T细胞的双荧光素酶活性。 -
用RIPA细胞裂解液从Hela细胞中提取蛋白质,并添加P8340蛋白酶抑制剂Cocktail进行 Western blot分析。随后在4℃下以13 500 r/min 离心15 min(离心半径为5 cm),收集上清液,并用BCA蛋白检测试剂盒进行定量。采用聚丙烯酰胺凝胶电泳(SDS-PAGE)(10%凝胶)分离蛋白质样品(20 μg)。将蛋白质转移至聚偏氟乙烯膜上,并在室温下用5% 牛血清白蛋白封闭2 h。温育后,使用抗UCHL5抗体、抗HIF-1α抗体、抗β-actin抗体、抗Bcl-2抗体在PBS中于4℃过夜孵育。将该膜与抗兔HRP标记的二抗在室温下孵育2 h(稀释比例为1∶1000)。采用增强的化学发光试剂进行显影,通过Quantity one软件系统拍照获得免疫印迹带。
-
从癌症基因组图谱(官方网站
https://portal.gdc.cancer.gov )中获得了宫颈癌和正常组织的基因表达数据,其中包括306个宫颈癌组织和13个宫颈正常组织。使用基因表达谱数据动态分析(官方网站http://gepia.cancer-pku.cn )进行基因相关性分析。 -
采用SPSS 24.0软件进行统计学分析。数据均符合正态分布、方差齐性检验后,组间差异比较采用单样本t检验;采用Pearson检验进行相关性分析。P<0.05为差异有统计学意义。
-
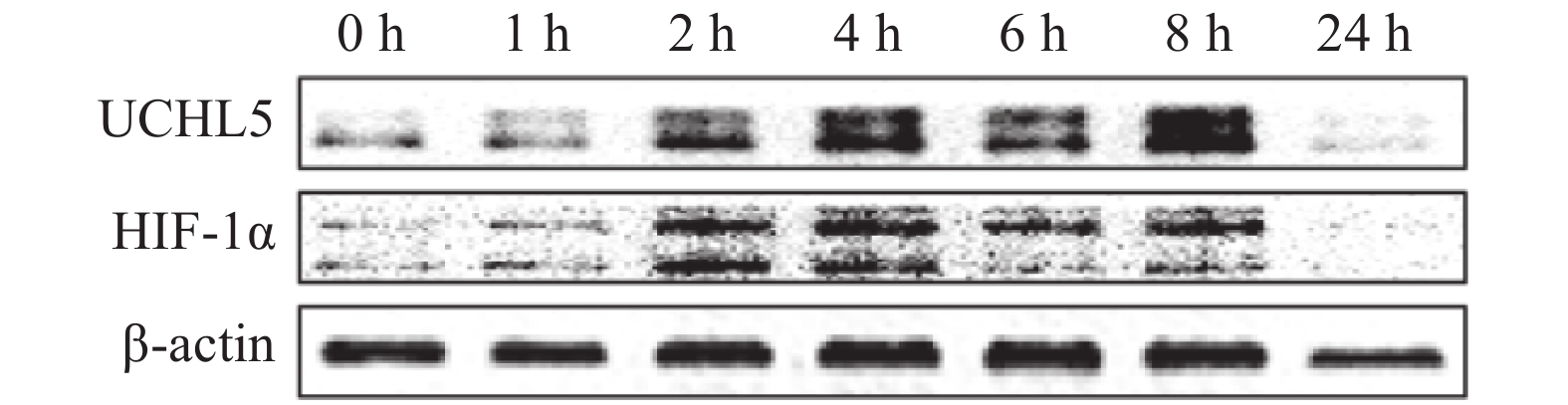
宫颈癌Hela细胞在缺氧条件下分别培养0、1、2、4、6、8、24 h后,观察发现UCHL5的表达水平出现上调,且在8 h时最明显(图1),这说明缺氧可诱导其表达。HIF-1α表达的改变趋势与UCHL5一致。

图 1 缺氧(1% O2)对宫颈癌 Hela 细胞 UHCL5 和 HIF-1α 的表达影响
Figure 1. Effect of hypoxia (1% O2) on the expression of ubiquitin carboxyl-terminal hydrolase isozyme L5 and hypoxia-inducible factor 1α in Hela cells of cervical cancer
-
Western blot和实时定量PCR结果显示,慢病毒载体感染Hela细胞后可显著上调或下调UCHL5的表达。其中,过表达UCHL5组转录本2和沉默UCHL5组转录本2的调控效率在蛋白和RNA水平上均较高(图2),所以选择此2种转录本进行后续实验。

图 2 上 调 ( A) 和 下 调 ( B)UCHL5后其蛋白的表达量和mRNA的相对含量
Figure 2. The protein expression and relative mRNA levels of up-regulated (A) and down-regulated (B) ubiquitin carboxyl-terminal hydrolase isozyme L5
-
图3的细胞克隆形成实验结果显示,与对照组(0 Gy)相比,分别给予2、4、6 Gy γ射线照射后,Hela细胞的克隆形成数目呈剂量依赖性减少(均P<0.05)。与过表达对照组相比,等剂量照射+过表达UCHL5组的细胞克隆形成率均增加,且差异有统计学意义(均P<0.05,图3A);与沉默对照组相比,等剂量照射+沉默UCHL5组的细胞克隆形成率均降低,且差异有统计学意义(均P<0.05,图3B)。

图 3 上调(A)和下调(B)UCHL5 在不同剂量 γ 射线照射后宫颈癌 Hela 细胞的克隆形成率
Figure 3. Colony formation rate of up-regulated (A) and down-regulated (B) ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels on cervical cancer Hela cells after different doses of γ-ray irradiation
-
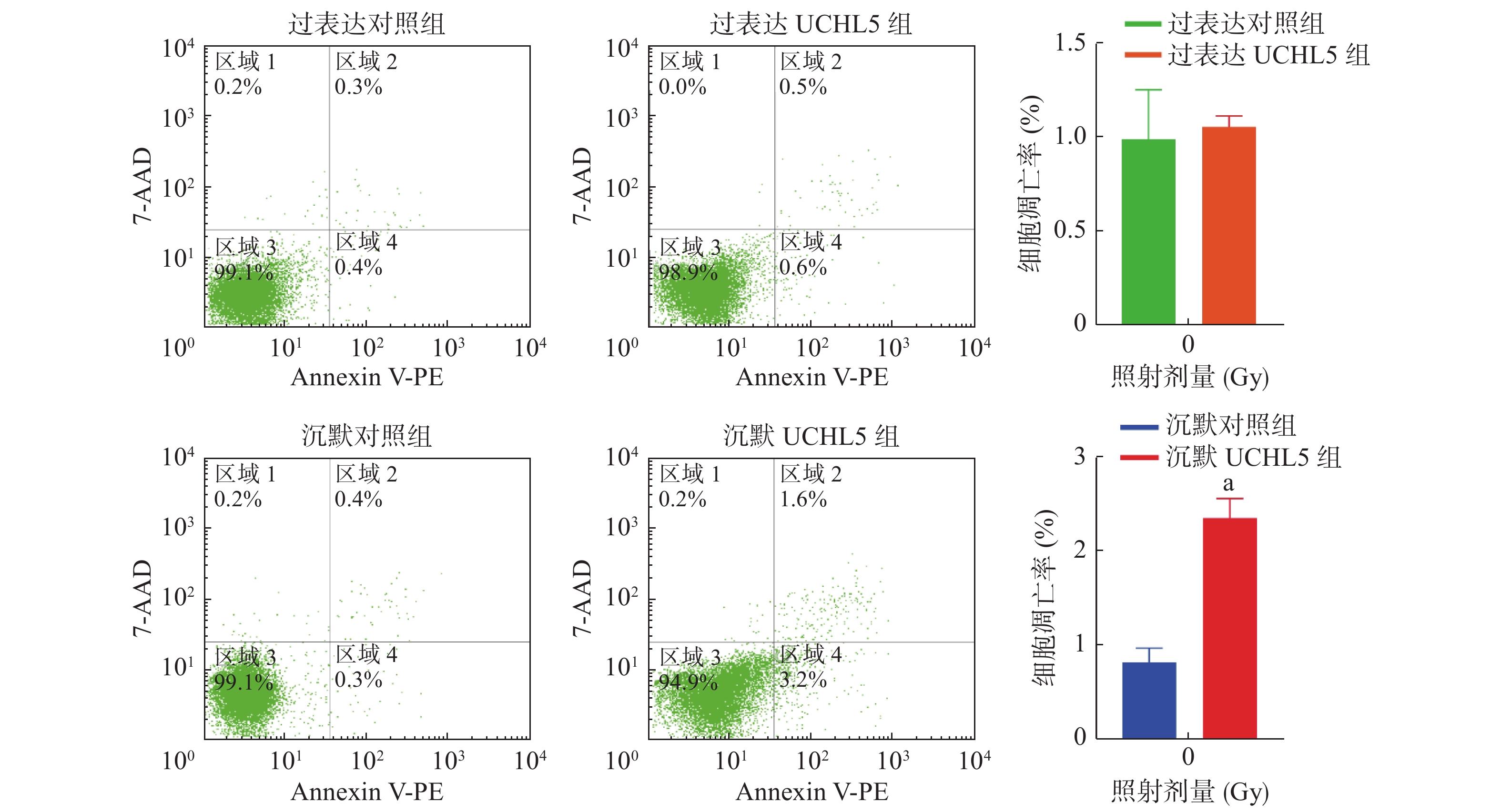
图4的流式细胞术检测结果显示,与沉默对照组相比,沉默UCHL5组的细胞凋亡率明显增加(P<0.05);与过表达对照组相比,过表达UCHL5组的细胞凋亡无明显变化,可能的原因是UCHL5在宫颈癌Hela细胞中表达的本底值较高,上调UCHL5表达后产生的效应不明显。以上结果表明,UCHL5低表达可促进宫颈癌Hela细胞的凋亡。

图 4 上调和下调UCHL5的宫颈癌 Hela 细胞凋亡率
Figure 4. Cell apoptosis rate of up-regulation and down-regulation of ubiquitin carboxyl-terminal hydrolase isozyme L5 in cervical cancer Hela cells
-
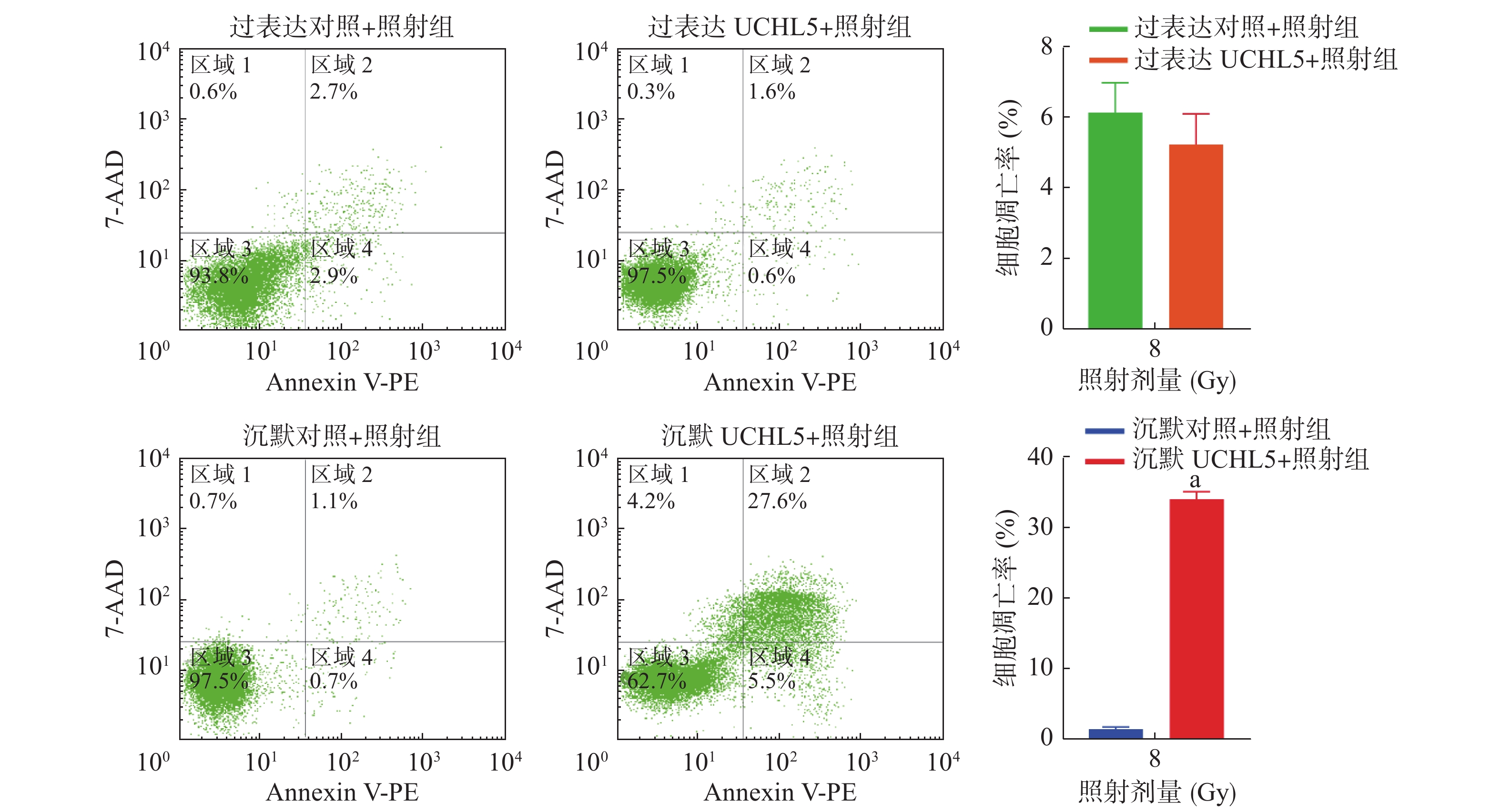
由图5可见,与沉默对照+照射组相比,沉默UCHL5+照射组宫颈癌Hela细胞的凋亡率明显增加,且差异有统计学意义(P<0.05);与过表达对照+照射组相比,过表达UCHL5+照射组宫颈癌Hela细胞的凋亡率下降,但差异无统计学意义(t=1.15,P>0.05)。这说明低表达UCHL5可以促进辐射诱导的细胞凋亡。

图 5 上调和下调UCHL5 在8 Gy γ 射线照射48 h后宫颈癌 Hela 细胞凋亡率
Figure 5. Up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 apoptosis rate of cervical cancer Hela cells after 8 Gy γ-ray irradiation 48 h
-
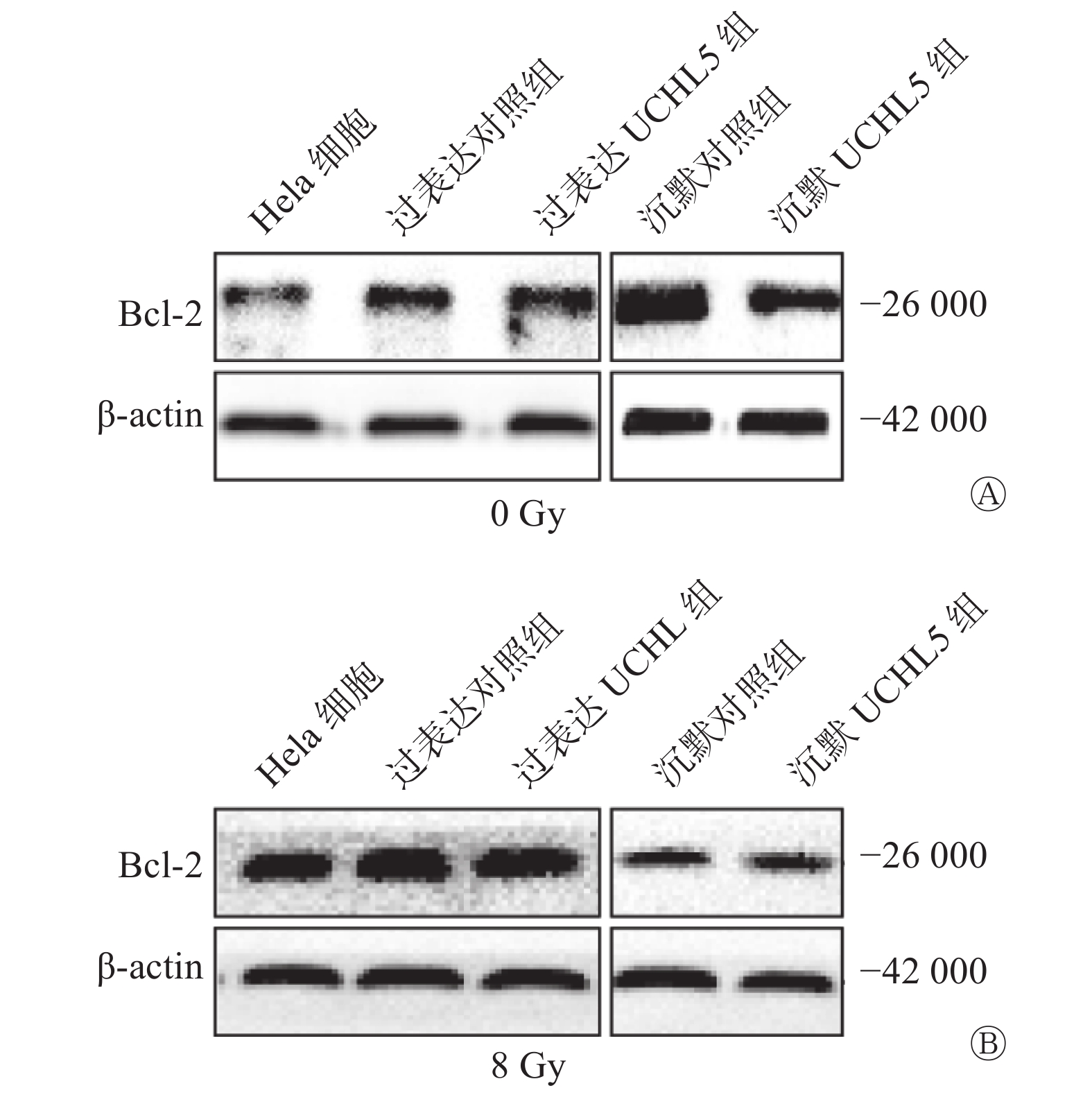
Western blot结果显示,过表达UCHL5组的Hela细胞中Bcl-2表达水平上升,沉默UCHL5组的Bcl-2表达水平下降(图6A)。8 Gy γ射线照射后48 h,过表达UCHL5组的Bcl-2表达水平相对于过表达对照组上升,沉默UCHL5组的Bcl-2表达水平相对于沉默对照组的变化不明显(图6B)。这说明UCHL5促进宫颈癌Hela细胞凋亡的发生可以通过促进抑制凋亡相关分子Bcl-2的表达水平而发挥作用。

图 6 上调和下调UCHL5 在8 Gy γ 射线照射前(A)、48 h后(B)宫颈癌 Hela 细胞Bcl-2 的表达量 UCHL5为泛素羧基末端水解酶L5;Bcl-2为B细胞淋巴瘤2;β-actin为β肌动蛋白
Figure 6. Bcl-2 expression levels before (A) and after (B) 48 h8 Gy γ-ray irradiation of up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels in cervical cancer Hela cells
-
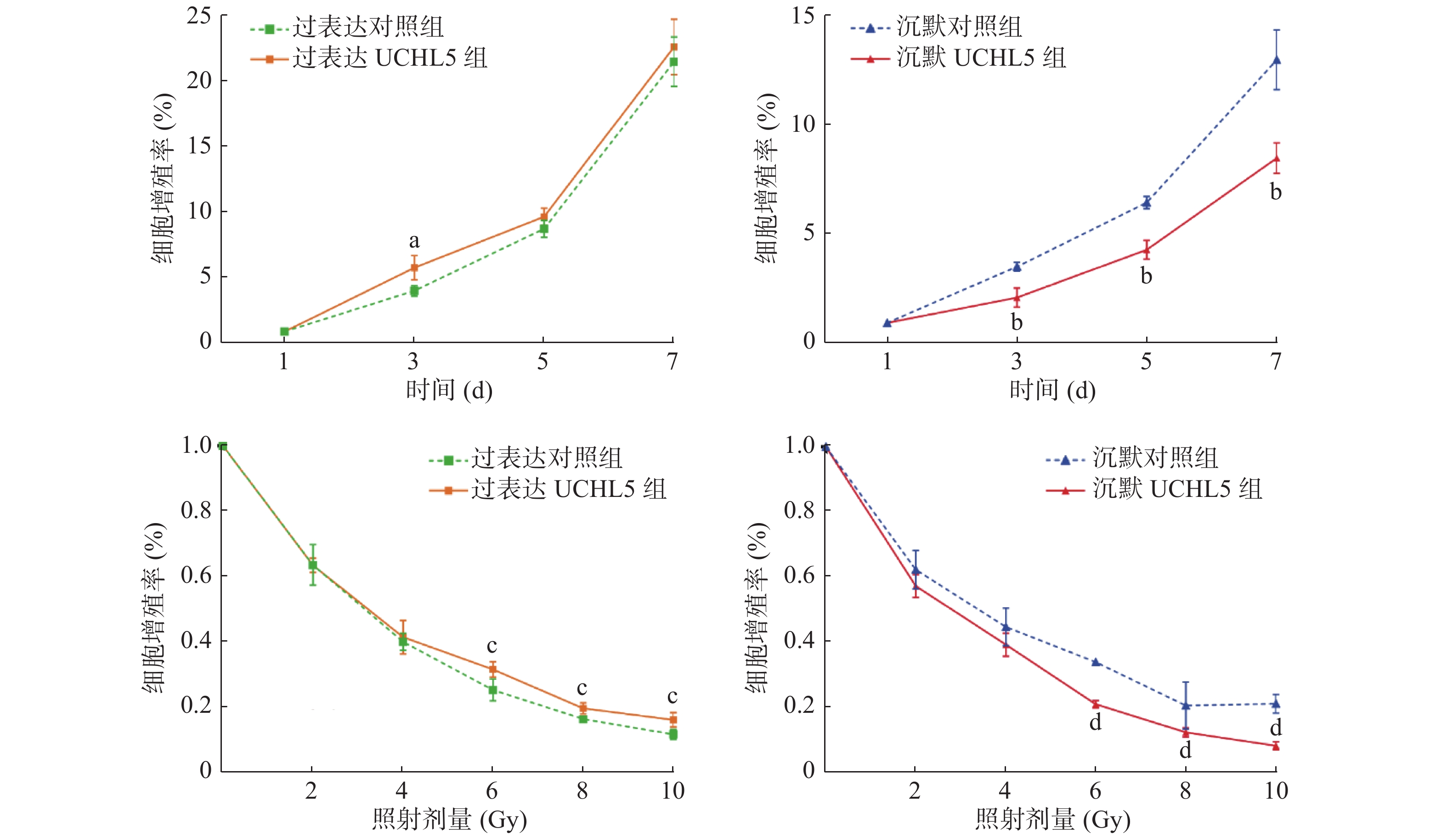
图7的MTT实验结果显示,与过表达对照组相比,过表达UCHL5组的Hela细胞增殖率较高,增殖率只有在第3天时的差异有统计学意义(P<0.05),这可能与UCHL5在Hela细胞中的表达的本底值较高有关;沉默UCHL5组的Hela细胞增殖率随着照射时间的延长均较沉默对照组降低,且差异有统计学意义(均 P<0.05)。与单纯照射组相比,给予不同剂量γ射线照射后,细胞的活力均呈剂量依赖性下降,过表达UCHL5组的Hela细胞的增殖率高于过表达对照组,且差异有统计学意义(均P<0.05);相反,沉默UCHL5组的细胞增殖率降低,且差异有统计学意义(均P<0.05)(图7),这表明降低UCHL5的表达可抑制Hela细胞的增殖。

图 7 上调和下调UCHL5 在不同时间或不同剂量 γ 射线照射后宫颈癌 Hela 细胞增殖率
Figure 7. Cell proliferation rate at different times or different doses of up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels in cervical cancer Hela cells
-
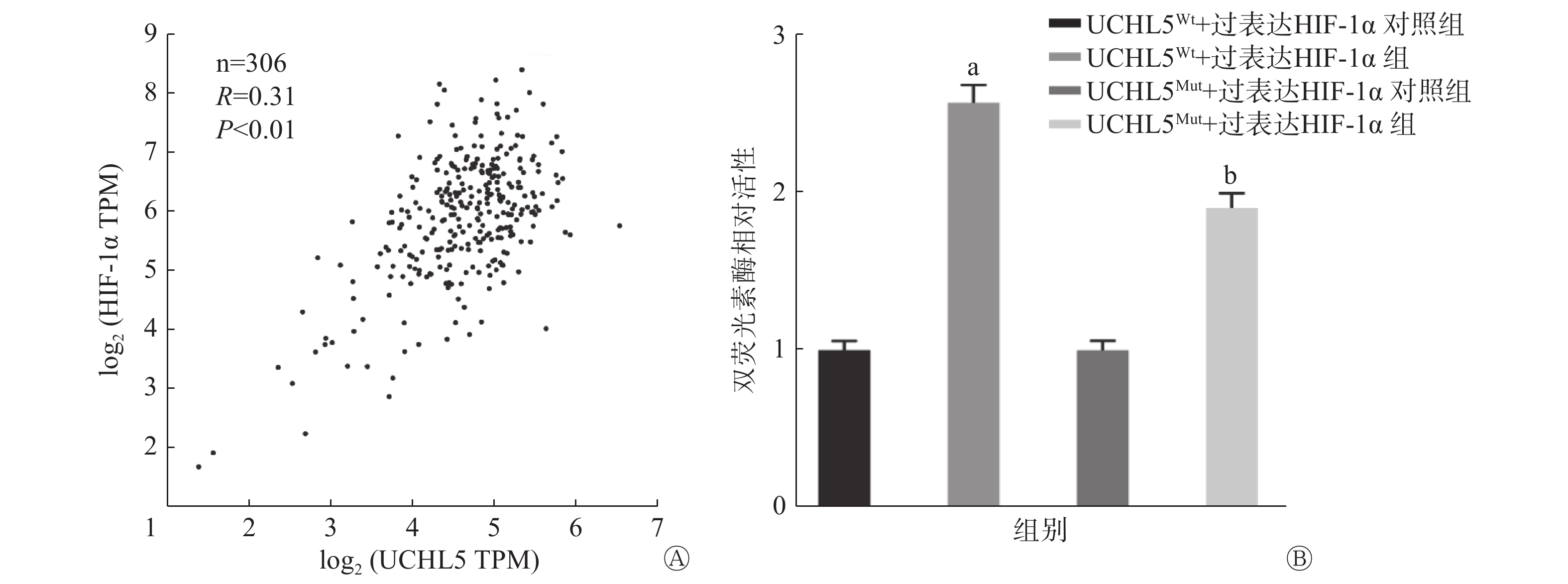
基因表达谱数据动态分析癌症基因组图谱数据显示,宫颈癌组织中HIF-1α的表达水平和UCHL5的表达水平呈正相关(P<0.01,图8A)。图8B的双荧光素酶报告基因结果表明,过表达HIF-1α可促进UCHL5启动子3′UTR(非翻译区)的荧光素酶活性,与UCHL5Wt+过表达HIF-1α对照组相比,UCHL5Wt+过表达HIF-1α组的活性增加了2.5倍,且差异有统计学意义(P<0.05),这说明HIF-1α可通过该结合位点调控UCHL5启动子的活性;而在结合位点突变后,UCHL5Mut+过表达HIF-1α组的活性高于UCHL5Mut+过表达HIF-1α对照组,且差异有统计学意义(P<0.05),这说明HIF-1α对UCHL5突变型的荧光素酶表达仍具有部分激活作用。以上结果表明,除突变的结合位点外,可能还存在其他非典型的结合位点。

图 8 宫颈癌 UCHL5和 HIF-1α 表达水平的相关性(A)、UCHL5在人肾上皮293T细胞中对HIF-1α的靶向调控作用(B)
Figure 8. The correlation between the expression levels of ubiquitin carboxyl-terminal hydrolase isozyme L5 and hypoxia-inducible factor 1α in cervical cancer tissues (A) and targeted regulation of ubiquitin carboxyl-terminal hydrolase isozyme L5 on hypoxia-inducible factor 1α in human renal epitheliar 293T cells (B)
-
宫颈癌是全球女性中第四大最常见的癌症,由于发展中国家宫颈癌的筛查普及率较低,大多数患者就诊时病情已进展至晚期或预后不良阶段[5]。目前,放疗仍是不可替代的治疗局部晚期宫颈癌的方法。随着科学技术的发展,放疗靶点的精确性不断提高[6]。但是,根治性放疗后宫颈癌患者的淋巴结复发率仍然高达10%~20%[7]。所以,辐射抗性细胞的存在仍是影响宫颈癌放疗疗效的重要因素。
实体瘤中普遍存在缺氧情况[8]。缺氧是影响肿瘤发生发展和治疗耐受性的重要原因。对于某些癌症,例如头颈鳞状细胞癌,缺氧是导致局部放疗失败的主要因素[9-10]。Gray等[11]首次发现,经高压氧处理后,荷瘤小鼠对辐射的敏感性显著增加,在氧合良好的条件下照射时,肿瘤细胞对X射线的灵敏度是缺氧条件下的3倍。我们课题组的前期研究结果表明,对Hela细胞进行缺氧预处理可以增强细胞的辐射抵抗能力,氧增强比为2.31±0.36[12]。
UCHL5在多种肿瘤中的表达上调,包括胃癌[13]、乳腺癌[14]、卵巢癌[15]、肝癌[16]、食管癌[17]以及宫颈癌[18],且提示预后不良。有研究报道,DUB在肿瘤辐射敏感性调节中发挥着重要作用:泛素特异性蛋白酶(ubiquitin-specific protease,USP)28通过稳定c-Myc/HIF-1α通路可以增强食管癌细胞的辐射抗性[19];卵巢肿瘤结构域蛋白4通过促进ATM(毛细血管扩张突变基因)/Chk2(细胞周期检测点激酶2)/p53信号通路和同源重组修复DNA双链损伤,增强非小细胞肺癌细胞的辐射抗性[20];USP38与组蛋白去乙酰化酶直接相互作用,稳定表达,促进组蛋白去乙酰化酶对组蛋白H3第56位氨基酸的乙酰化修饰,增强了非同源末端的连接效率,促进了肿瘤细胞的辐射抗性[21]。目前,关于UCHL5对肿瘤辐射敏感性影响的研究未见报道。缺氧作为应激源可以调节DUB,比如缺氧时,肺癌A549细胞的USP28表达下调[22];USP46在缺氧条件下的表达下调,导致结肠癌中肿瘤抑制因子PHLPP丝氨酸/苏氨酸蛋白磷酸酶的降解,使得多种结肠癌细胞系的化疗耐药性增加[23];在缺氧条件下,USP47去泛素化β链蛋白也被诱导表达,进而增强直肠癌细胞上皮间质转化[24-25]。本研究结果显示,缺氧诱导宫颈癌Hela细胞UCHL5的表达,提示该基因产物可能是缺氧影响宫颈癌肿瘤辐射敏感性的重要因子。此外,低表达UCHL5降低了Hela细胞的增殖能力,增加了细胞的辐射敏感性,促进了Hela细胞的凋亡以及γ射线诱导的凋亡。这表明UCHL5可以促进宫颈癌的辐射抗性,并且通过促进细胞增殖和抑制细胞凋亡发挥作用。有研究报道,HIFs直接参与缺氧介导的UCHL1过表达,HIF-1α和HIF-2α可以结合UCHL1启动子中潜在的缺氧反应元件(HRE)位点,导致UCHL1的转录增加,从而促进缺氧诱导的神经元损伤细胞的凋亡[26]。但是,HIF-1α对UCHL5的调控作用尚未见报道,本研究结果显示,宫颈癌组织中HIF-1α的表达与UCHL5有关,在非氧气依赖的条件下,HIF-1α对UCHL5的转录有激活作用,这表明UCHL5在缺氧条件下表达水平升高可能是通过HIF-1α转录激活的方式实现的。
近年来,DUB成为热门的癌症治疗靶标,如果简单地认为促进低氧性癌症中HIF-α的降解是有益的,那么使用DUB抑制剂拮抗E3泛素连接酶对HIF-α的降解将具有重要的临床意义。然而,连接酶E3和DUB也受缺氧的调节,使得这一作用机制变得复杂。此外,DUB的作用不仅限于缺氧信号传导,还存在于其他信号传导途径,如DNA损伤修复、细胞周期调控等。所以,UCHL5在HIF信号通路之内或之外以及两者之间的联系需要进一步探索。
综上所述,本研究证实了缺氧诱导宫颈癌Hela细胞UCHL5的表达;过表达UCHL5能增强细胞的增殖能力,抑制细胞凋亡,增强细胞的辐射抗性。在生物机制上,缺氧诱导的HIF-1α可以激活UCHL5转录。因此,靶向缺氧诱导的UCHL5的特异性抑制剂可能成为一种潜在的放疗增敏剂,但缺氧诱导UCHL5的表达进而影响宫颈癌辐射敏感性的机制还需深入研究。
利益冲突 本研究由署名作者按以下贡献声明独立开展,不涉及任何利益冲突。
作者贡献声明 焦圣元、高巧慧负责实验数据的整理分析、实验的实施和论文的撰写;达飞、郭利协助实验实施;王晋、郭娟、李静参与实验的设计;刘军叶负责指导课题的设计和论文的审阅。
缺氧诱导的UCHL5增强宫颈癌Hela细胞辐射抗性
Hypoxia-induced UCHL5 expression enhanced radioresistance of Hela cells in cervical cancer
-
摘要:
目的 研究缺氧诱导的泛素羧基末端水解酶L5(UCHL5)在调节宫颈癌Hela细胞辐射敏感性中的作用。 方法 以宫颈癌Hela细胞为研究对象,1% O2条件下培养观察UCHL5表达水平的变化。采用Western blot和实时定量聚合酶链反应(PCR)检测慢病毒载体感染Hela细胞后稳定调变UCHL5的效率,转录本分为空白对照组、过表达对照组、过表达UCHL5组转录本1~4和沉默对照组、沉默UCHL5组转录本1~2。将实验所用的细胞分为过表达对照组、过表达UCHL5组、沉默对照组和沉默UCHL5组。采用流式细胞术检测8 Gy γ射线照射48 h后细胞的凋亡率;采用细胞克隆形成实验检测0、2、4、6 Gy γ射线单次照射培养2周后4组细胞的克隆形成率;采用3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT)实验检测4组细胞培养1周后的增殖率以及联合0、2、4、6、8、10 Gy γ射线照射后的增殖率。使用基因表达谱数据动态分析癌症基因组图谱数据库宫颈癌组织和正常组织中抗缺氧诱导因子1α(HIF-1α)与UCHL5表达的相关性。采用双荧光素酶报告基因实验观察HIF-1α对UCHL5的激活作用。组间比较采用单样本t检验,采用Pearson检验进行相关性分析。 结果 缺氧可诱导宫颈癌Hela细胞UCHL5的表达。Western blot和实时定量PCR结果显示,感染后的Hela细胞可显著上调或下调UCHL5的表达,其中,过表达UCHL5组转录本2和沉默UCHL5组转录本2的表达均较高,所以选择此2种转录本进行后续实验。克隆形成实验结果显示,与接受相同剂量照射的过表达对照组相比,上调UCHL5增加了Hela细胞的克隆形成率,在0 、2、4、6 Gy剂量照射后克隆形成率的差异均有统计学意义( t=14.16、19.22、8.76、6.79,均 P<0.05)。 流式细胞术结果显示,与沉默对照组相比,沉默 UCHL5 促进了 Hela 细胞的凋亡(t=10.29,P<0.05),增加了 γ 射线诱导的细胞凋亡率 (t=52.01, P<0.05)。MTT 实验结果显示,与过表达对照组相比,上调 UCHL5 可升高宫颈癌 Hela 细胞的增殖率,增殖率在第3天时差异有统计学意义 (t=3.905,P<0.05);与过表达对照组相比,等剂量照射 UCHL5 上调组升高了受照细胞的增殖率,在剂量为6、8、10 Gy时,细胞的增殖率差异均有统计学意义(t=3.40、4.06、3.68,均P<0.05)。宫颈癌组织中 HIF-1α 表达水平和UCHL5 表达水平呈正相关(R=0.31,P<0.01),双荧光素酶报告基因结果显示 HIF-1α 结合并激活 UCHL5 启动子的活性,其活性增加了2.5倍(t=30.47,P<0.05)。 结论 缺氧条件下,宫颈癌Hela细胞中UHCL5的诱导表达降低了细胞的辐射敏感性,其潜在的机制可能与HIF-1α转录激活UCHL5的表达有关。 -
关键词:
- 低氧 /
- 宫颈肿瘤 /
- 辐射抗性 /
- 去泛素化酶 /
- 泛素羧基末端水解酶 L5
Abstract:Objective To investigate the role of hypoxia-induced ubiquitin carboxyl-terminal hydrolase isozyme L5 (UCHL5) in the regulation of radiosensitivity of Hela cells in cervical cancer. Methods UCHL5 levels were detected in Hela cells cultured under 1% O2 condition. Western blot and qRT-PCR analyses verified the efficiency of lentiviral vector infection on stable UCHL5 modulation in HeLa cells, including oe-vector, oe-UCHL5-1, oe-UCHL5-2, oe-UCHL5-3, oe-UCHL5-4 and sh-vector, sh-UCHL5-1 and sh-UCHL5-2. The cells used in the experiment were divided into the following groups: oe-vector, oe-UCHL5, sh-vector and sh-UCHL5. Cell colony-formation rate and radiosensitivity were detected by colony-formation assay combined with single-dose (0, 2, 4 and 6 Gy) γ-ray irradiation after culturing for 2 weeks. The effect of radiation on cell apoptosis was determined by flow cytometry after 48 h of 8 Gy γ-ray irradiation. 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay was used to detect the cell viability of down- or up-regulated UCHL5 cells before and after 0, 2, 4, 6, 8 and 10 Gy of γ-ray irradiation. Gene expression profiling interactive analysis was used to analyze the correlated expression between UCHL5 and hypoxia inducible factor-1α (HIF-1α). The transcriptional activation effect of UCHL5 by HIF-1α was detected using dual-luciferase reporter-gene assay. Differences between groups were compared by single-sample t test, and Pearson test was used for correlation analysis. Results UCHL5 was significantly induced by hypoxia after culturing at different time points. Hela cell lines with stably overexpressed and silenced UCHL5 were successfully constructed, with oe-UCHL5-2 and sh-UCHL5-2 having the highest regulatory efficiency. These two groups were selected for subsequent experiments. Compared with the control group that received the same dose of irradiation, significant differences existed at doses of 0, 2, 4 and 6 Gy (t=14.16, 19.22, 8.76, 6.79, all P<0.05), respectively. Knock-down of UCHL5 promoted apoptosis (t=10.29, P<0.05) and radiation-induced apoptosis (t=52.01, P<0.05). UCHL5 up-regulation promoted cell proliferation, and the proliferation rate was statistically significant on the third day (t=3.905, P<0.05). Furthermore, UCHL5 strengthened the viability (t=3.40, 4.06, 3.68, all P<0.05) of irradiated Hela cells, with significant differences at doses of 6, 8 and 10 Gy (t=3.40, 4.06, 3.68, all P<0.05), respectively. The expression levels of HIF-1α and UCHL5 in cervical cancer tissues were positively correlated (R=0.31, P<0.01). Additionally, HIF-1α was a potential transcriptional activator of UCHL5 in Hela cells, and its activity increased 2.5 times (t=30.47, P<0.05). Conclusions The induced expression of UHCL5 in cervical cancer Hela cells under hypoxia condition can reduce the radiation sensitivity of cells. The underlying mechanism may be related to the HIF-1α transcriptional activation of UCHL5 expression. -
图 1 缺氧(1% O2)对宫颈癌 Hela 细胞 UHCL5 和 HIF-1α 的表达影响
Figure 1. Effect of hypoxia (1% O2) on the expression of ubiquitin carboxyl-terminal hydrolase isozyme L5 and hypoxia-inducible factor 1α in Hela cells of cervical cancer
图 2 上 调 ( A) 和 下 调 ( B)UCHL5后其蛋白的表达量和mRNA的相对含量
Figure 2. The protein expression and relative mRNA levels of up-regulated (A) and down-regulated (B) ubiquitin carboxyl-terminal hydrolase isozyme L5
图 3 上调(A)和下调(B)UCHL5 在不同剂量 γ 射线照射后宫颈癌 Hela 细胞的克隆形成率
Figure 3. Colony formation rate of up-regulated (A) and down-regulated (B) ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels on cervical cancer Hela cells after different doses of γ-ray irradiation
图 4 上调和下调UCHL5的宫颈癌 Hela 细胞凋亡率
Figure 4. Cell apoptosis rate of up-regulation and down-regulation of ubiquitin carboxyl-terminal hydrolase isozyme L5 in cervical cancer Hela cells
图 5 上调和下调UCHL5 在8 Gy γ 射线照射48 h后宫颈癌 Hela 细胞凋亡率
Figure 5. Up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 apoptosis rate of cervical cancer Hela cells after 8 Gy γ-ray irradiation 48 h
图 6 上调和下调UCHL5 在8 Gy γ 射线照射前(A)、48 h后(B)宫颈癌 Hela 细胞Bcl-2 的表达量 UCHL5为泛素羧基末端水解酶L5;Bcl-2为B细胞淋巴瘤2;β-actin为β肌动蛋白
Figure 6. Bcl-2 expression levels before (A) and after (B) 48 h8 Gy γ-ray irradiation of up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels in cervical cancer Hela cells
图 7 上调和下调UCHL5 在不同时间或不同剂量 γ 射线照射后宫颈癌 Hela 细胞增殖率
Figure 7. Cell proliferation rate at different times or different doses of up-regulated and down-regulated ubiquitin carboxyl-terminal hydrolase isozyme L5 expression levels in cervical cancer Hela cells
图 8 宫颈癌 UCHL5和 HIF-1α 表达水平的相关性(A)、UCHL5在人肾上皮293T细胞中对HIF-1α的靶向调控作用(B)
Figure 8. The correlation between the expression levels of ubiquitin carboxyl-terminal hydrolase isozyme L5 and hypoxia-inducible factor 1α in cervical cancer tissues (A) and targeted regulation of ubiquitin carboxyl-terminal hydrolase isozyme L5 on hypoxia-inducible factor 1α in human renal epitheliar 293T cells (B)
-
[1] Shrestha AD, Neupane D, Vedsted P, et al. Cervical cancer prevalence, incidence and mortality in low and middle income countries: a systematic review[J]. Asian Pac J Cancer Prev, 2018, 19(2): 319−324. DOI: 10.22034/APJCP.2018.19.2.319. [2] Nishi R, Wijnhoven P, Le Sage C, et al. Systematic characterization of deubiquitylating enzymes for roles in maintaining genome integrity[J]. Nat Cell Biol, 2014, 16(10): 1016−1026. DOI: 10.1038/ncb3028. [3] Randles L, Anchoori PK, Roden RBS, et al. The proteasome ubiquitin receptor hRpn13 and its interacting deubiquitinating enzyme uch37 are required for proper cell cycle progression[J]. J Biol Chem, 2016, 291(16): 8773−8783. DOI: 10.1074/jbc.M115.694588. [4] Chen ZW, Niu XM, Li ZM, et al. Effect of ubiquitin carboxy-terminal hydrolase 37 on apoptotic in A549 cells[J]. Cell Biochem Funct, 2011, 29(2): 142−148. DOI: 10.1002/cbf.1734. [5] Lee H, Kang Y, Ju W. Cervical cancer screening in developing countries: using visual inspection methods[J]. Clin J Oncol Nurs, 2016, 20(1): 79−83. DOI: 10.1188/16.CJON.79-83. [6] Vordermark D. Radiotherapy of cervical cancer[J]. Oncol Res Treat, 2016, 39(9): 516−520. DOI: 10.1159/000448902. [7] Huang EY, Wang CJ, Chen HC, et al. Multivariate analysis of para-aortic lymph node recurrence after definitive radiotherapy for stage IB-IVA squamous cell carcinoma of uterine cervix[J]. Int J Radiat Oncol Biol Phys, 2008, 72(3): 834−842. DOI: 10.1016/j.ijrobp.2008.01.035. [8] Nussenbaum F, Herman IM. Tumor angiogenesis: insights and innovations[J]. J Oncol, 2010, 2010: 132641. DOI: 10.1155/2010/132641. [9] Suh YE, Raulf N, Gäken J, et al. MicroRNA-196a promotes an oncogenic effect in head and neck cancer cells by suppressing annexin A1 and enhancing radioresistance[J]. Int J Cancer, 2015, 137(5): 1021−1034. DOI: 10.1002/ijc.29397. [10] Alsahafi E, Begg K, Amelio I, et al. Clinical update on head and neck cancer: molecular biology and ongoing challenges[J/OL]. Cell Death Dis, 2019, 10(8): 540[2020-10-21]. https://www.nature.com/articles/s41419-019-1769-9. DOI: 10.1038/s41419-019-1769-9. [11] Gray LH, Conger AD, Ebert M, et al. The concentration of oxygen dissolved in tissues at the time of irradiation as a factor in radiotherapy[J]. Br J Radiol, 1953, 26(312): 638−648. DOI: 10.1259/0007-1285-26-312-638. [12] Liu JY, Zhang J, Wang XW, et al. HIF-1 and NDRG2 contribute to hypoxia-induced radioresistance of cervical cancer Hela cells[J]. Exp Cell Res, 2010, 316(12): 1985−1993. DOI: 10.1016/j.yexcr.2010.02.028. [13] Arpalahti L, Laitinen A, Hagström J, et al. Positive cytoplasmic UCHL5 tumor expression in gastric cancer is linked to improved prognosis[J/OL]. PLoS One, 2018, 13(2): e0193125[2020-10-21]. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0193125. DOI: 10.1371/journal.pone.0193125. [14] Xia XH, Liao YN, Guo ZQ, et al. Targeting proteasome-associated deubiquitinases as a novel strategy for the treatment of estrogen receptor-positive breast cancer[J/OL]. Oncogenesis, 2018, 7(9): 75[2020-10-21]. https://www.nature.com/articles/s41389-018-0086-y. DOI: 10.1038/s41389-018-0086-y. [15] Wang L, Chen YJ, Xu K, et al. High expression of UCH37 is significantly associated with poor prognosis in human epithelial ovarian cancer[J]. Tumour Biol, 2014, 35(11): 11427−11433. DOI: 10.1007/s13277-014-2446-3. [16] Fang Y, Fu D, Tang WQ, et al. Ubiquitin C-terminal hydrolase 37, a novel predictor for hepatocellular carcinoma recurrence, promotes cell migration and invasion via interacting and deubiquitinating PRP19[J]. Biochim Biophys Acta, 2013, 1833(3): 559−572. DOI: 10.1016/j.bbamcr.2012.11.020. [17] Chen YJ, Fu D, Xi JJ, et al. Expression and clinical significance of UCH37 in human esophageal squamous cell carcinoma[J]. Dig Dis Sci, 2012, 57(9): 2310−2317. DOI: 10.1007/s10620-012-2181-9. [18] Rolén U, Kobzeva V, Gasparjan N, et al. Activity profiling of deubiquitinating enzymes in cervical carcinoma biopsies and cell lines[J]. Mol Carcinog, 2006, 45(4): 260−269. DOI: 10.1002/mc.20177. [19] Zhang WL, Liu ZK, Wang JM, et al. Knockdown of USP28 enhances the radiosensitivity of esophageal cancer cells via the c-Myc/hypoxia-inducible factor-1 alpha pathway[J]. J Cell Biochem, 2019, 120(1): 201−212. DOI: 10.1002/jcb.27305. [20] Wu ZQ, Qiu MH, Guo Y, et al. OTU deubiquitinase 4 is silenced and radiosensitizes non-small cell lung cancer cells via inhibiting DNA repair[J/OL]. Cancer Cell Int, 2019, 19: 99[2020-10-21]. https://cancerci.biomedcentral.com/articles/10.1186/s12935-019-0816-z. DOI: 10.1186/s12935-019-0816-z. [21] Yang YF, Yang CZ, Li TT, et al. The deubiquitinase USP38 promotes NHEJ repair through regulation of HDAC1 activity and regulates cancer cell response to genotoxic insults[J]. Cancer Res, 2020, 80(4): 719−731. DOI: 10.1158/0008-5472.CAN-19-2149. [22] Li Q, Kluz T, Sun H, et al. Mechanisms of c-Myc degradation by nickel compounds and hypoxia[J/OL]. PLoS One, 2009, 4(12): e8531[2020-10-21]. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0008531. DOI: 10.1371/journal.pone.0008531. [23] Wen YA, Stevens PD, Gasser ML, et al. Downregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cells[J]. Mol Cell Biol, 2013, 33(22): 4594−4605. DOI: 10.1128/MCB.00695-13. [24] Shi JD, Liu YJ, Xu XH, et al. Deubiquitinase USP47/UBP64E regulates β-catenin ubiquitination and degradation and plays a positive role in wnt signaling[J]. Mol Cell Biol, 2015, 35(19): 3301−3311. DOI: 10.1128/MCB.00373-15. [25] Choi BJ, Park SA, Lee SY, et al. Hypoxia induces epithelial-mesenchymal transition in colorectal cancer cells through ubiquitin-specific protease 47-mediated stabilization of Snail: A potential role of Sox9[J/OL]. Sci Rep, 2017, 7(1): 15918[2020-10-21]. https://www.nature.com/articles/s41598-017-15139-5. DOI: 10.1038/s41598-017-15139-5. [26] Wu HW, Ying W, Wang W, et al. HIF1α and HIF2α mediated UCHL1 upregulation in hypoxia-induced neuronal injury following neuronal hypoxic ischemic encephalopathy[J]. Int J Clin Exp Pathol, 2016, 9(2): 2677−2685. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3924
- HTML全文浏览量: 2868
- PDF下载量: 30