


 下载:
下载:

-
目前,放疗仍是肺癌、乳腺癌和食道癌等胸部恶性肿瘤患者的重要治疗手段[1]。肺是对辐射中等敏感的器官,放疗会导致肿瘤周围的正常肺组织受到不同程度的损伤。近年来,随着对放射性肺损伤机制认识的逐步加深,人们渐渐发现免疫学机制也是其发生和发展的一个重要原因[2]。在电离辐射的作用下,肺组织中免疫细胞的分布和特性发生改变,进而参与放射性肺损伤。其中,调节性T细胞(regulatory T cells,Treg)是一种重要的免疫调节细胞,属于CD4+T细胞的亚群,有研究报道,Treg可通过分泌转化生长因子β(transforming growth factor-beta,TGF-β)等细胞因子来调节各种原因引起的肺部炎症和肺纤维化[3-5]。
Fontenot等[6]研究结果证实,叉头样转录因子3(forkhead transcription factor 3,Foxp3)在CD4+ CD25+Treg中特异性表达,并且是CD4+ CD25+Treg发育和功能发挥的关键调节因子。因此,Foxp3被认为是Treg的特异性标志物。还有大量研究结果表明,除了Foxp3外,神经纤毛蛋白1(neuropilin 1,NRP1)是Treg的另一种表面标志物,胸腺来源的天然型Treg表面高表达NRP1,而外周来源的诱导型Treg表面NRP1的表达量很低,这两种细胞均表现出相似的免疫抑制活性及控制稳态条件下自身免疫反应的作用[7-9]。但这两种细胞在功能上的差异一直是学者们关注的焦点。
T淋巴细胞在各种损伤引起的炎症反应中发挥着错综复杂的作用,不同类型的T细胞通过分泌不同的细胞因子来调节免疫反应的平衡。在放射性肺损伤中,Treg与T细胞的其他亚群,如辅助T细胞(helper T cell,Th),包括Th17、Th1和Th2型细胞等相互关联,各自分泌的代表性细胞因子[TGF-β、白细胞介素(interleukin,IL)-17A、干扰素γ(interferon-γ,IFN-γ)、IL-2和IL-4)]更是参与其中,形成复杂的调节网络,影响放射性肺损伤的发生发展。
目前,已有研究结果表明Treg参与放射性肺纤维化的发生与发展[10],那么Treg作为免疫调节细胞是否参与放射性肺损伤的炎症阶段?Treg的分化对放射性肺损伤的发展起什么作用?本研究将围绕以上问题,深入探究Treg的分化对放射性肺损伤的影响及其作用机制。
-
纯化免疫球蛋白G(immunoglobulin G,IgG)抗体和CD25抗体(PC61)购自美国Biolegend公司;异硫氰酸荧光素(FITC)-CD4抗体、别藻蓝蛋白(APC)-CD25抗体、藻红蛋白(phycoerythrin,PE)-Foxp3抗体和Foxp3固定破膜液购自美国BD Biosciences 公司;PE-CD25抗体、TGF-β1、IL-17A、IFN-γ、IL-2和IL-4酶联免疫吸附测定试剂盒购自美国Thermo Fisher公司;NRP1抗体购自美国Abcam公司;Ⅰ型胶原酶购自美国GIBCO公司;Masson三色染色试剂盒购自福州迈新生物技术有限公司;抗原修复液、山羊血清购自北京索莱宝科技有限公司;抗荧光淬灭剂购自上海翊圣生物科技有限公司;二喹啉甲酸(BCA)蛋白浓度测定试剂盒、蛋白裂解液(RIPA Lysis Buffer)和蛋白酶抑制剂(苯甲基磺酰氟)购自上海碧云天生物技术有限公司;绿色荧光二抗[异硫氰酸荧光素(FITC)-兔抗小鼠抗体]购自美国Bioworld公司。
-
清洁级健康C57BL/6小鼠,雌性,6~8周龄,体重(20±2)g,共48只,购自北京华阜康生物科技股份有限公司,合格证号:SCXK(京)2014-0004。小鼠饲养于吉林大学基础医学院实验动物中心,饲养间温度为(23±2)℃,每12 h明暗交替照明。
-
采用X射线辐照仪(X-RAD 320iX,美国PXI公司)对小鼠进行单次全胸照射(其余部位用铅板遮挡),照射剂量为20 Gy,剂量率为2.22 Gy/min,电压320 V,电流12.5 mA,源皮距60 cm。
-
将C57BL/6小鼠按随机数字表法分成4组:空白对照组、单纯照射组、照射+IgG组和照射+CD25组,每组12只,采用1.3节中的照射条件对小鼠进行照射,照射+IgG组和照射+CD25组于照射后2 h分别腹腔注射IgG抗体和CD25抗体各100 μg,每周1次,并于照射后第4周和第8周各处死小鼠6只。采用流式细胞术检测各组小鼠肺组织中CD25+Foxp3+Treg的百分比来鉴定小鼠Treg抑制模型是否建立成功。
-
将各组小鼠肺组织制备成单细胞悬液,用PBS洗2次,重悬于1%牛血清白蛋白中,加入5 μL 别藻蓝蛋白(APC)-CD25抗体,于4℃避光孵育45 min,PBS洗涤后加入固定破膜液1 mL,孵育45 min,用PBS洗2次,加入5 μL PE-Foxp3抗体,于4℃避光孵育45 min,用PBS洗2次,加入500 μL的3%牛血清白蛋白,随后上流式细胞仪(BD FACSCalibur,美国BD公司)进行检测。
-
将照射后第4周和第8周的小鼠肺组织进行研磨,每组0.1 g,加入400 μL的蛋白裂解液和4 μL蛋白酶抑制剂,提取细胞总蛋白,采用二喹啉甲酸(BCA)蛋白浓度测定试剂盒测定蛋白浓度,10%十二烷基磺酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳100 min,4℃转膜75 min,室温下封闭1 h,一抗4℃孵育过夜,TBST(Tris缓冲液-Tween-20)洗涤,二抗常温孵育1 h,TBST洗涤,用电化学发光(ECL)液显色。采用ImageJ软件(
https://imagej.net/Welcome )测定各显色条带的灰度值,计算目的蛋白条带与甘油醛-3-磷酸脱氢酶蛋白条带(GAPDH)(内参蛋白)灰度值的比值,即目的蛋白表达量,并以空白对照组的蛋白表达量为1,计算其余各组蛋白的相对表达量。 -
将石蜡切片脱蜡至水,置于抗原修复液中进行抗原修复,用200 μL的5%山羊血清进行封闭,一抗用50 μL的NRP1抗体(1∶250稀释)和PE-CD25抗体(1∶250稀释)共同孵育,于湿盒内4℃过夜,滴加绿色荧光二抗(1∶500稀释)进行孵育,用抗荧光淬灭剂封片,于荧光显微镜下观察。
-
采用5%水合氯醛(0.1 mL/10 g)对各组小鼠进行麻醉,麻醉后分别拍摄每只小鼠腹面和背面的照片,观察小鼠皮肤的损伤程度。
-
将各组小鼠麻醉后,从右心室进针,冲出肺内血液,分离肺脏,置于4%多聚甲醛中固定24 h,流水冲洗24 h后将组织包埋,制作石蜡切片,行复水、苏木精-伊红染色和脱水处理。
-
取照射后第4周和第8周的小鼠肺组织,每组0.1 g,加入400 μL PBS,用组织研磨仪进行研磨,离心半径8 cm,3000 r/min离心5 min,取上清保存于−80℃冰箱中,按照酶联免疫吸附测定试剂盒的说明书步骤,分别检测肺组织中Treg代表性细胞因子TGF-β1和各种亚型Th细胞代表性细胞因子(Th17型细胞代表性细胞因子IL-17A、Th1型细胞代表性细胞因子IFN-γ和IL-2、Th2型细胞代表性细胞因子IL-4)的分泌水平。
-
采用SPSS 24.0软件进行统计学分析。实验数据符合正态分布且方差齐,以均数±标准差(
$ \bar x \pm s$ )表示,两组间比较采用独立样本t检验。P<0.05为差异有统计学意义。 -
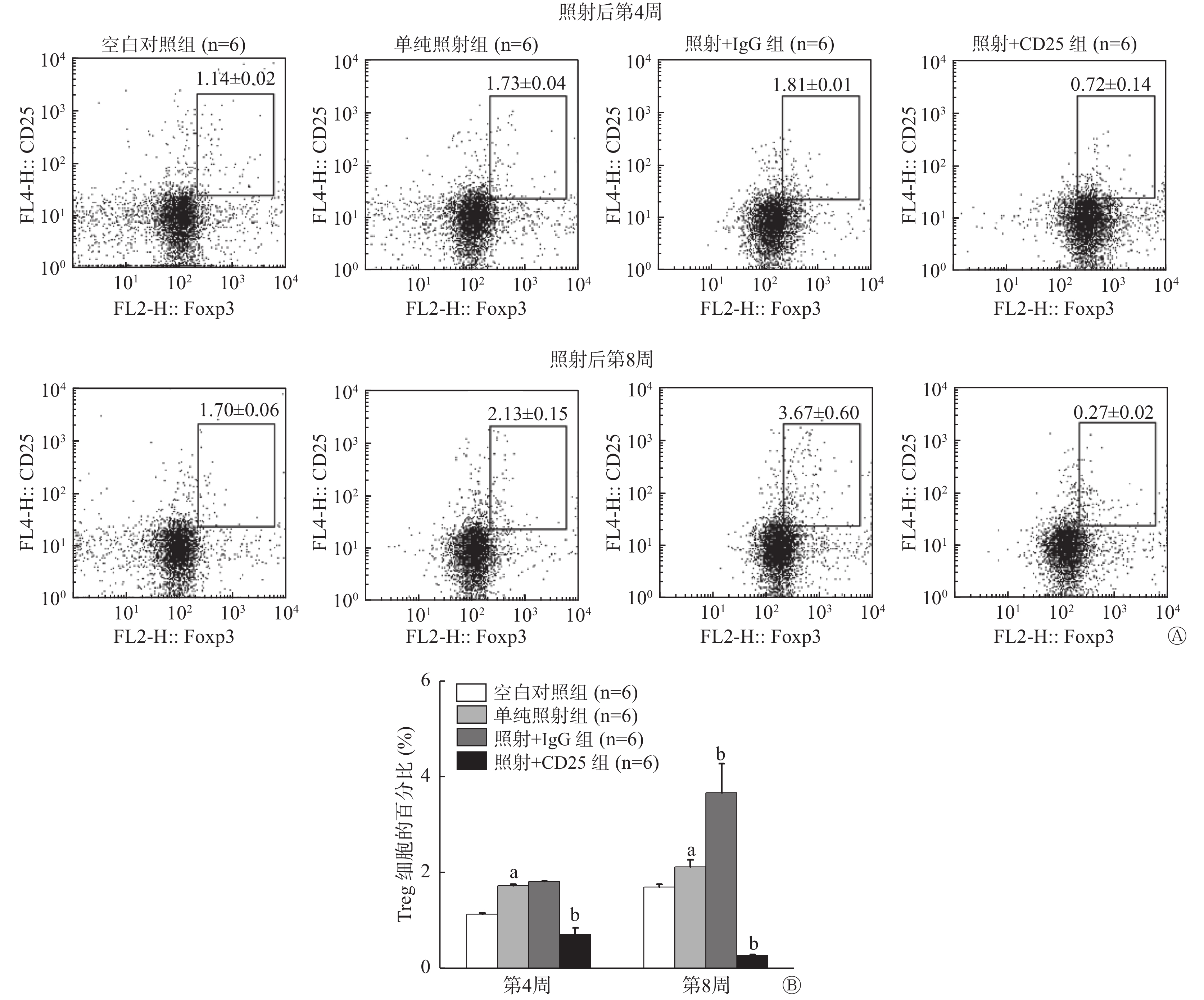
流式细胞术分析结果显示,照射后第4周和第8周,单纯照射组小鼠肺组织内CD25+Foxp3+Treg百分比分别为(1.73±0.04)%和(2.13±0.15)%,均较空白对照组[(1.14±0.02)% 和 (1.70±0.06)%] 明显升高,且差异均有统计学意义(t=−26.680、−4.545,P=0.000、0.010)。照射+IgG组小鼠肺组织内CD25+Foxp3+Treg百分比在照射后第4周上升至(1.81±0.04)%,第8周上升至(3.67±0.60)%,与单纯照射组相比,差异有统计学意义(t=−5.667,P=0.012)。照射+CD25组小鼠肺组织内CD25+Foxp3+Treg的百分比于照射后第4周下降至(0.72±0.14)%,与单纯照射组相比,差异有统计学意义(t=5.296,P=0.000),照射后第8周其下降幅度更大,降至(0.27±0.02)%,与单纯照射组相比,差异有统计学意义(t=37.538,P=0.000)(图1)。此结果说明我们构建的小鼠Treg抑制模型建立成功。
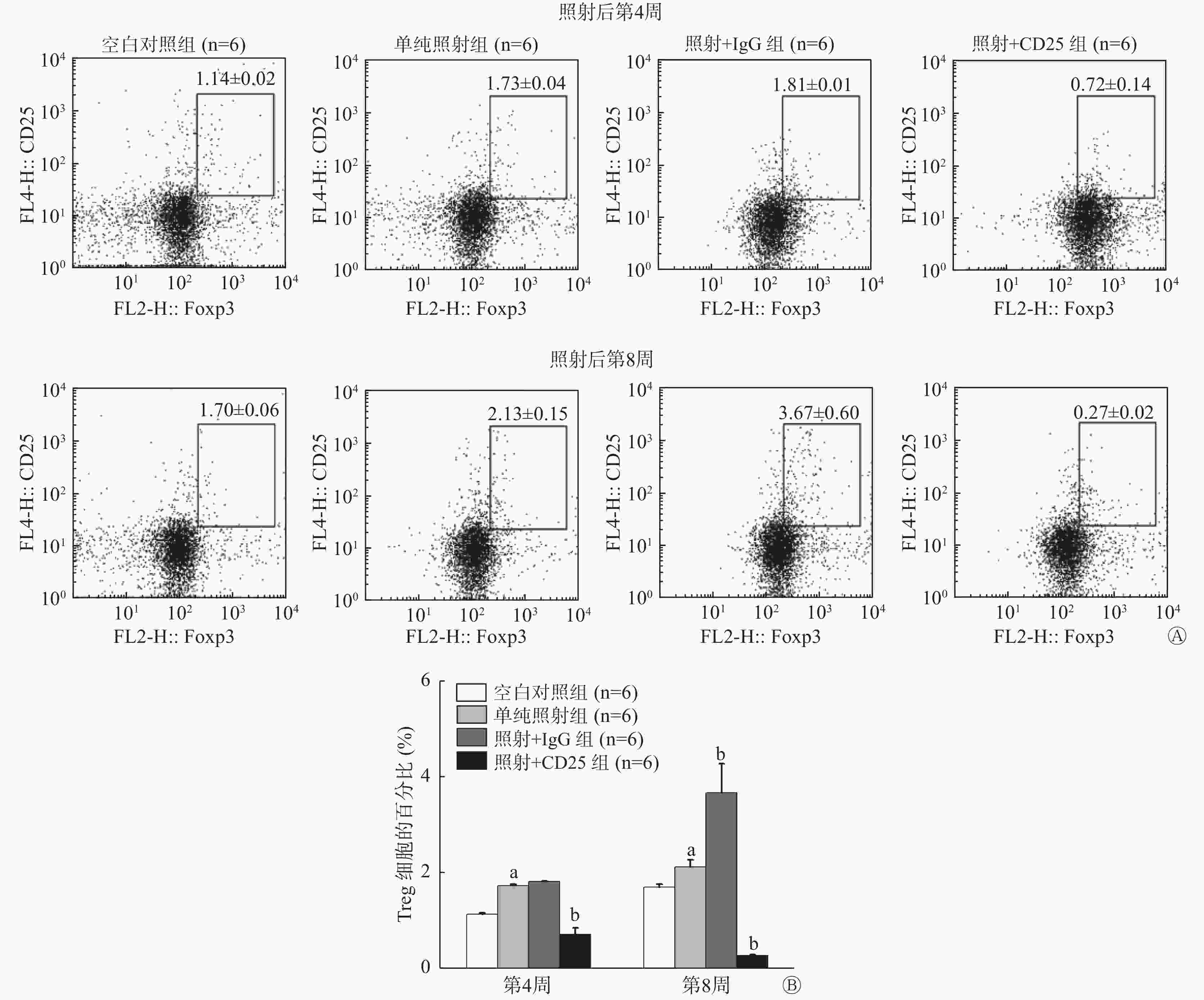
图 1 20 Gy X射线照射后C57BL/6小鼠Treg抑制模型肺组织中CD25+Foxp3+Treg的百分比
Figure 1. Percentage of CD25+Foxp3+Tregs in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
-
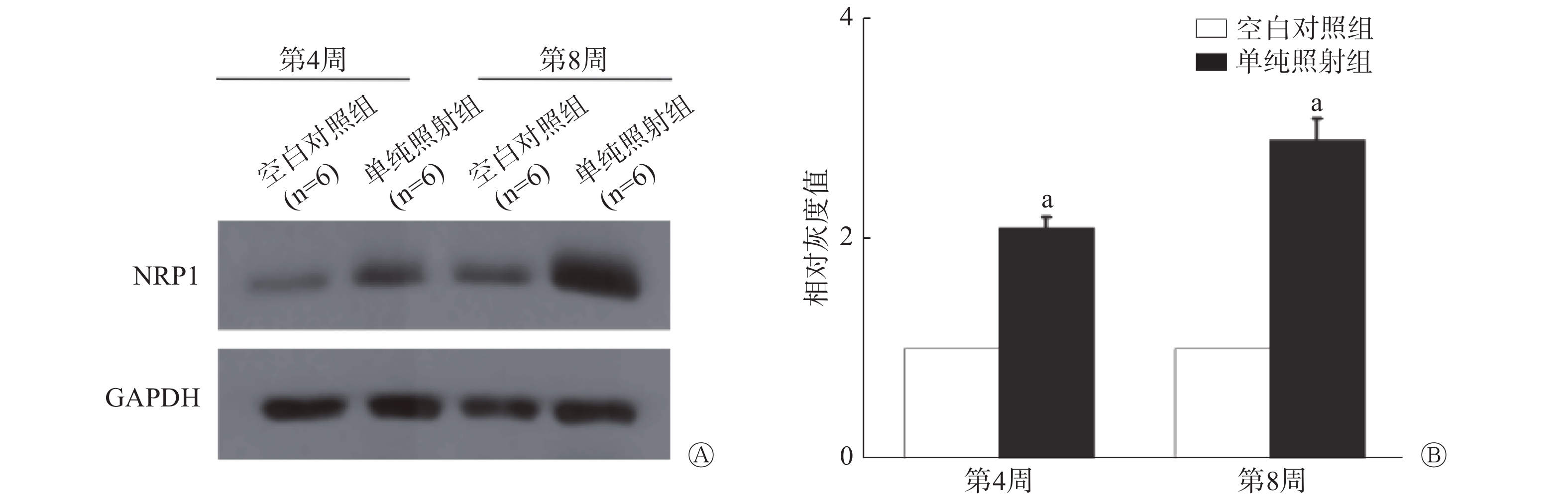
如图2所示,照射后第4周,单纯照射组小鼠肺组织内NRP1蛋白的表达量是空白对照组的2.06倍(t=−7.341,P=0.000),照射后第8周,NRP1蛋白的表达量是空白对照组的2.90倍(t=−9.127,P=0.000)。此结果提示NRP1可能参与放射性肺损伤的发生。
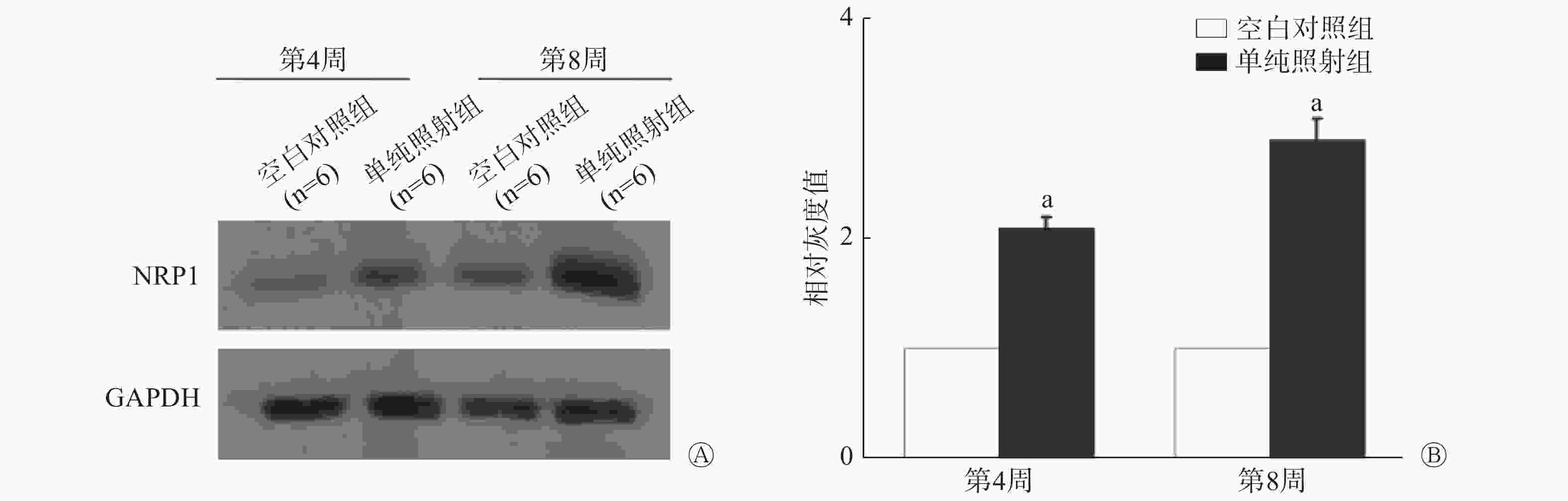
图 2 20 Gy X 射线照射后C57BL/6小鼠肺组织内NRP1蛋白的表达量变化
Figure 2. Changes of NRP1 protein in lung tissue of the C57BL/6 mouse irradiated by 20 Gy X-ray
-
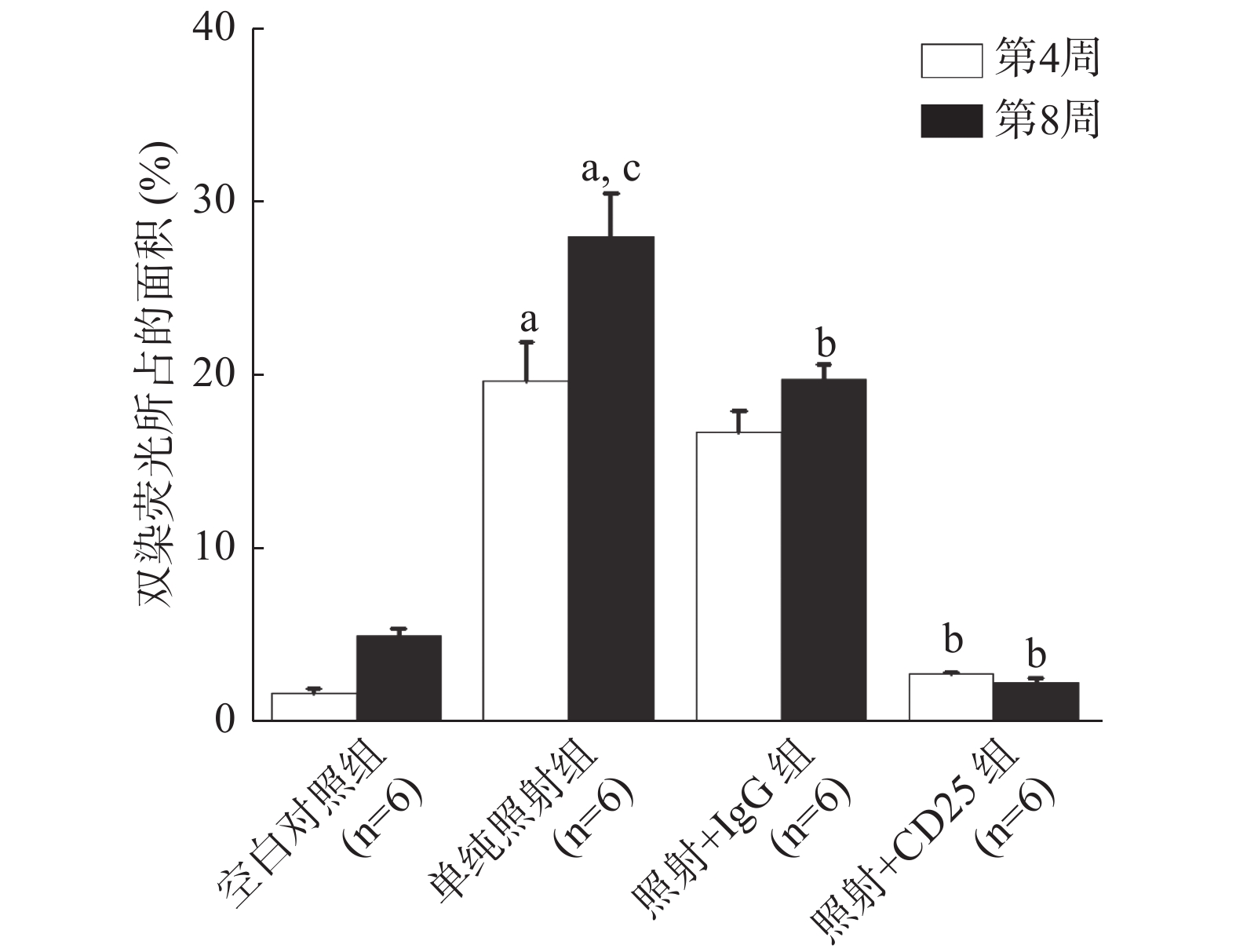
与空白对照组相比,照射后第4周,单纯照射组和照射+IgG组NRP1和CD25共同标记的Treg的荧光面积比例(即:CD25+NRP1+Treg百分比)均增加,这说明照射后CD25+NRP1+Treg的分化增多,照射后第8周这种变化更加显著。而照射后第4周和第8周,照射+CD25组中CD25+NRP1+Treg的荧光面积比例较单纯照射组降低,且差异均有统计学意义(t=8.926、14.457,P=0.001、0.000)(图3)。

图 3 20 Gy X 射线照射后C57BL/6小鼠Treg抑制模型肺组织内CD25+NRP1+Treg的百分比
Figure 3. Percentage of CD25+NRP1+Tregs in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
-
观察小鼠皮肤损伤情况后发现,空白对照组小鼠皮肤正常,但照射后第4周单纯照射组和照射+IgG组小鼠胸廓照射野皮肤出现明显的放射性损伤,表现为脱毛、水肿以及炎性渗出;而照射+CD25组皮肤损伤明显减轻。照射后第8周单纯照射组和照射+IgG组由第4周时的脱毛、水肿发展为脱屑、溃疡,损伤逐渐加重,而照射+CD25组也逐渐出现皮肤损伤,表现为脱毛和干性脱皮,但损伤程度远低于单纯照射组和照射+IgG组(图4中A)。病理学结果显示,照射后第4周,空白对照组小鼠肺组织未见异常改变,即肺组织结构清晰,肺泡壁完整;单纯照射组和照射+IgG组小鼠肺组织损伤较为严重,镜下均可见肺泡炎性改变(少量的炎性细胞如中性粒细胞浸润),肺泡壁增厚,肺间质水肿,肺间质可见毛细血管扩张;而照射+CD25组小鼠肺组织结构清晰,肺泡壁未见增厚,这表明Treg受抑制后可减轻放射性肺损伤。照射后第8周,单纯照射组和照射+IgG组小鼠肺组织损伤程度较照射后第4周更为严重,肺泡壁出现大范围破损、断裂,大量炎性细胞浸润,液体渗出(图4中B)。
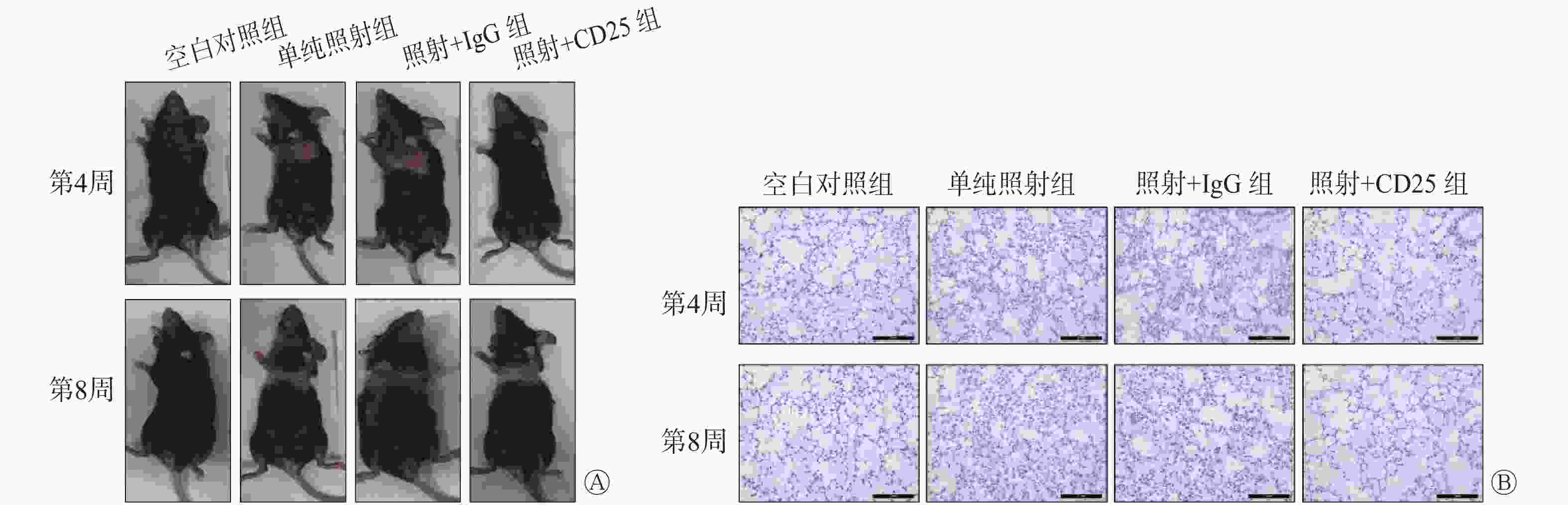
图 4 20 Gy X 射线照射后C57BL/6小鼠Treg抑制模型中各组小鼠的皮肤损伤及肺组织的病理学改变 图中,A:皮肤损伤的变化;B:肺组织的病理学变化(苏木精-伊红染色,×20)。IgG:免疫球蛋白G;Treg:调节性T细胞
Figure 4. Skin damage and pathological changes in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
-
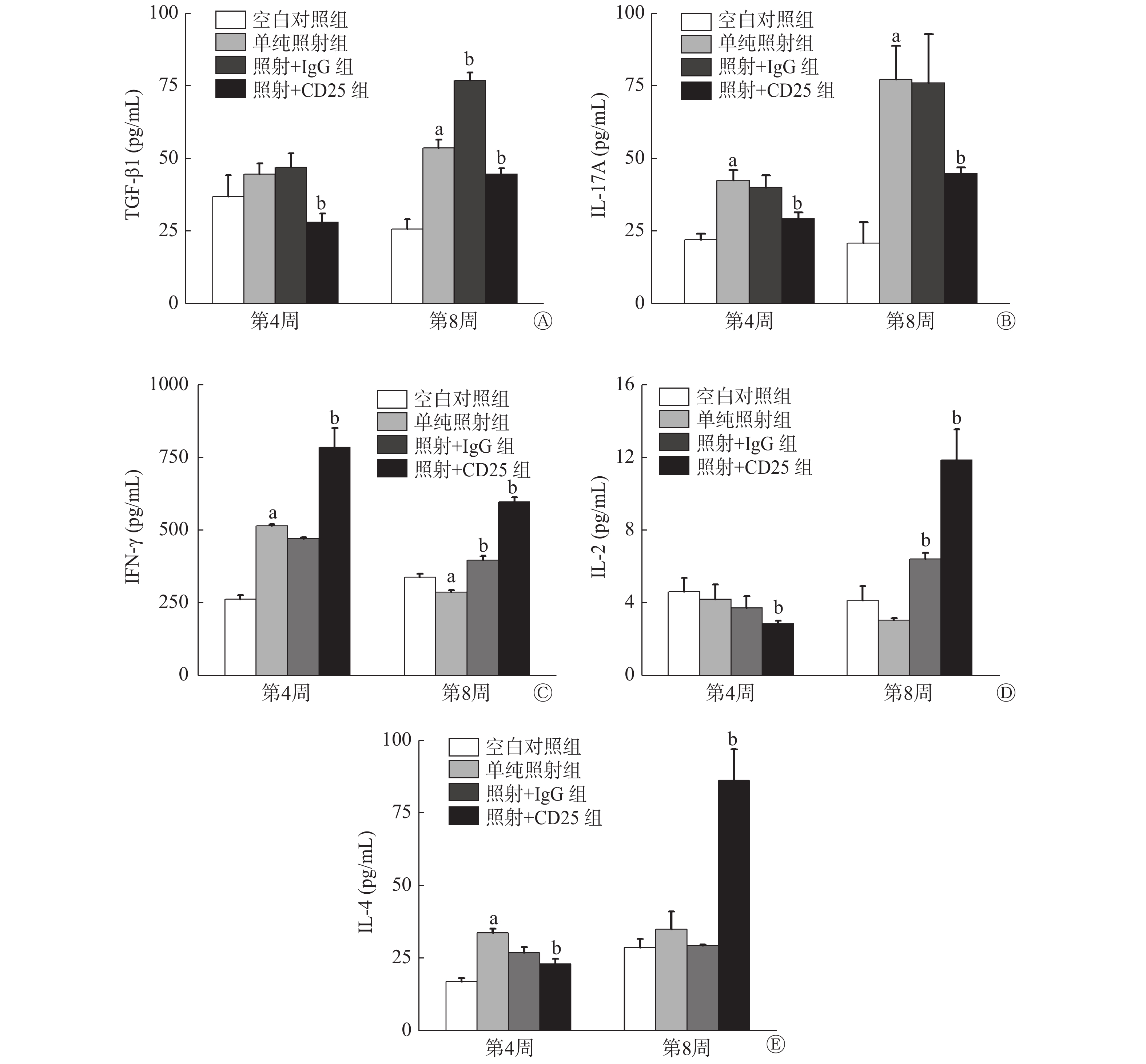
Treg分泌的代表性细胞因子的变化情况:与空白对照组相比,单纯照射组TGF-β1水平在照射后第4周升高,而且第8周升高更明显(t=−11.072,P=0.000);与单纯照射组相比,照射+CD25组TGF-β1水平在照射后第4周和第8周均降低(t=6.037、4.524,均 P=0.000),这说明抑制Treg可以抑制TGF-β1的分泌(图5中A)。
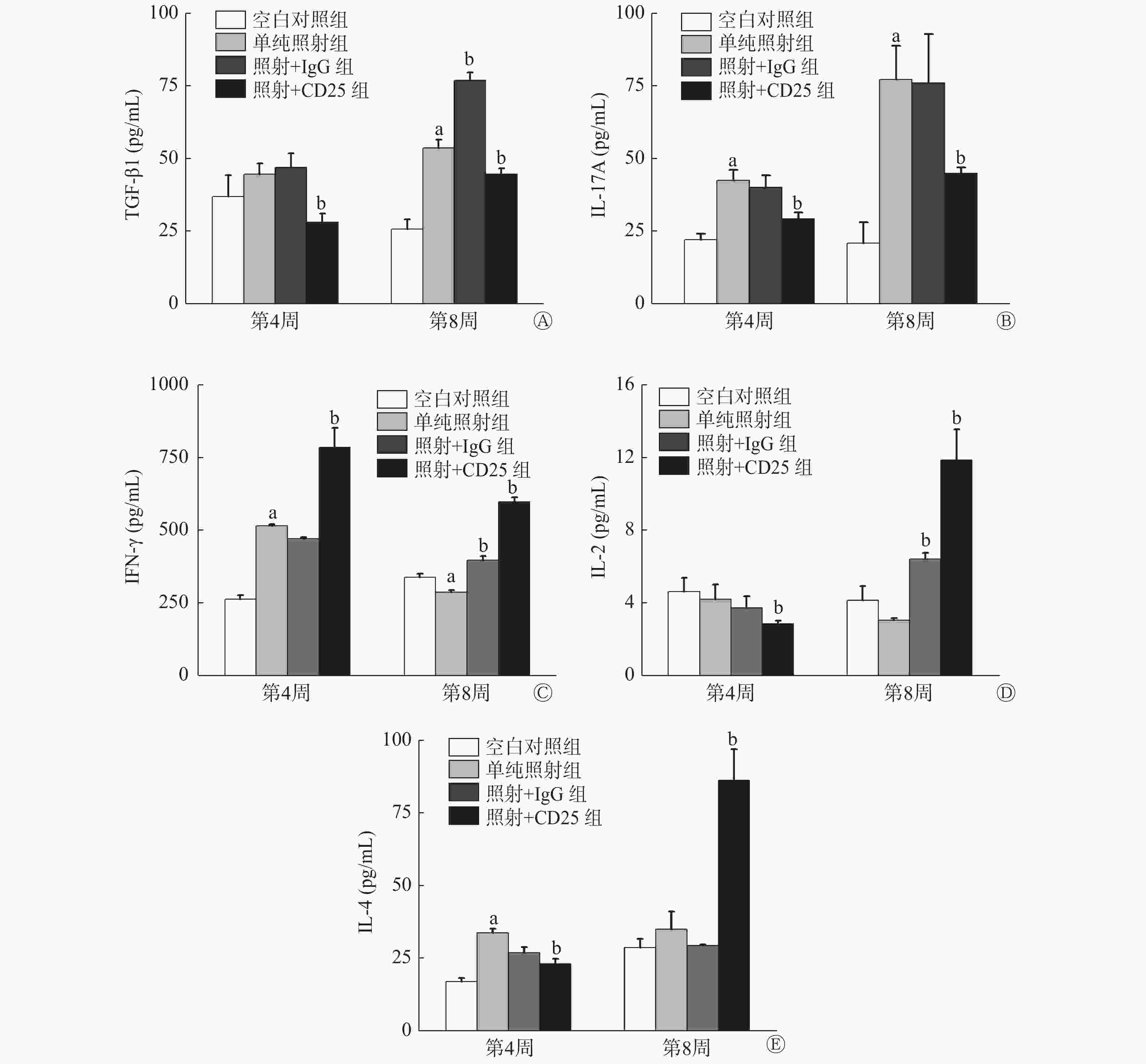
图 5 20 Gy X 射线照射后C57BL/6小鼠Treg抑制模型肺组织中分泌的相关细胞因子的水平变化
Figure 5. Secretion of related cytokines in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
Th17型细胞分泌的代表性细胞因子的变化情况:与空白对照组相比,单纯照射组的IL-17A水平在照射后第4周和第8周明显升高( t=−8.492、−7.167, P=0.001、0.002);与单纯照射组相比,照射+CD25组的IL-17A水平在第4周和第8周均降低( t=5.496、4.772,P=0.000),且在照射后第8周降低更明显,但较空白对照组仍是升高的,说明辐射诱导的肺损伤中IL-17A明显升高,促进炎症,而抑制了Treg后,IL-17A水平降低,炎症程度也有所减轻(图5中B)。
Th1型细胞分泌的代表性细胞因子的变化情况:与空白对照组相比,单纯照射组的IFN-γ在照射后第4周升高( t=−27.393, P=0.000),第8周时除了单纯照射组IFN-γ水平略有降低外,照射+IgG组和照射+CD25组IFN-γ均升高;与单纯照射组相比,照射+CD25组的IFN-γ水平在照射后第4周和第8周均明显升高(t=−7.006、−12.565, P=0.002、0.000)(图5中C);与空白对照组相比,单纯照射组、照射+IgG组和照射+CD25组的IL-2水平在照射后第4周均降低,而在照射后第8周,照射+CD25组的IL-2水平明显比单纯照射组高出约2倍(t=−9.090, P=0.000)(图5中D)。
Th2型细胞分泌的代表性细胞因子的变化情况:与空白对照组相比,单纯照射组的IL-4水平在照射后第4周( t=−15.796, P=0.000)和第8周均升高;与单纯照射组相比,照射+CD25组的IL-4水平在照射后第4周降低( t=8.833, P=0.000),第8周明显升高(t=−7.191, P=0.000)(图5中E)。
-
CD4+CD25+Foxp3+Treg在维持机体免疫稳态和耐受中起着关键作用,其通过分泌各种细胞因子(如IL-10和TGF-β等)或抑制效应T细胞等方式来维持体内免疫系统的平衡[11]。其中,胸腺来源的天然型Treg(NRP1hiTreg)和外周来源的诱导型Treg(NRP1lowTreg)都能在IL-12表达的基础上,上调Th1相关分子的表达,包括T-box基因家族转录因子(T-bet)、趋化因子受体3(CXCR3)和IFN-γ。有文献报道,天然型Treg在炎症条件下不稳定,可能在IL-6存在下转变为Th17表型[12]。相反,诱导型Treg在体外和体内炎症条件下比较稳定。在胶原诱导的关节炎小鼠模型中,诱导型Treg相对于天然型Treg对破骨细胞生成和骨侵蚀显示出更好的抑制作用[13]。但是,诱导型Treg在维持外周耐受中的确切作用仍不十分清楚,需要进一步探讨。
我们为了了解Treg分化在放射性肺损伤的形成过程中的作用,通过每周腹腔注射CD25单克隆抗体建立了抑制Treg的小鼠模型[14-16]。流式细胞术结果表明,照射+CD25组小鼠肺组织内CD25+Foxp3+Treg的百分比较单纯照射组降低,这说明Treg抑制模型建立成功。之后,我们发现单纯照射组小鼠肺组织内NRP1蛋白表达量升高,并进一步通过免疫荧光实验,结果发现照射后小鼠肺组织内CD25+NRP1+Treg增多,同时CD25+Foxp3+Treg(总Treg)也增多,所以我们推测CD25+NRP1+Treg和CD25+Foxp3+Treg均参与了放射性肺损伤的形成过程。Wirsdörfer等[17]也进行过类似的研究,他们发现肺组织在照射早期的肺炎症阶段会引发CD4+Foxp3+Treg的局部和全身积聚,这与我们的研究结果一致。从小鼠皮肤损伤的表现可以看出,单纯照射后的小鼠皮肤发生脱毛、溃烂和出血,但是照射后注射CD25抗体的小鼠皮肤的脱毛和糜烂现象明显改善,这说明抑制Treg可以明显缓解辐射对小鼠皮肤造成的放射性损伤。随着照射后时间的推移,单纯照射组和照射+IgG组在照射后8周,小鼠皮肤损伤逐渐加重,且照射+CD25组也逐渐出现皮肤损伤,我们推测,在放射性损伤的初期,注射CD25抗体可起到抑制损伤的作用,照射后8周这种作用依然存在,但由于损伤的逐步发展,CD25抗体的作用被抵消,从而小鼠皮肤表现出放射性损伤症状。从病理学结果可以看出,照射后第4周,单纯照射组和照射+IgG组小鼠肺组织发生炎症反应,第8周后损伤加重。而抑制Treg后小鼠肺组织结构清晰,没有出现炎性改变,这表明Treg受抑制后可减轻放射性肺损伤。
小鼠Treg抑制模型建立成功后,我们检测了肺组织内CD25+NRP1+Treg的表达变化,结果发现照射+CD25组小鼠肺组织中CD25+NRP1+Treg百分比相对于单纯照射组下降,由此我们认为CD25+NRP1+Treg与CD25+Foxp3+Treg发挥着相似的功能,可以促进早期的放射性肺损伤。
肺组织受照后发生放射性肺损伤,主要表现为放射性肺炎,发生炎症后就会有大量的炎性细胞聚集,本研究结果显示,Treg在受照后的肺组织中聚集,且伴随着促炎因子TGF-β1增多,而在小鼠Treg抑制模型中,被招募到受损肺部的Treg明显减少,且肺组织中TGF-β1减少,这说明Treg参与了放射性肺损伤的过程,且当Treg抑制时放射性肺损伤减轻。小鼠皮肤损伤程度的结果也证实了我们的这一观点。已有文献证明,Thl型细胞因子IFN-γ在其他因素诱导的肺纤维化中具有抑制成纤维细胞增殖的作用,即抑制纤维化的功能[18]。本研究结果证明,照射后给予CD25抗体抑制Treg后,IFN-γ增多,发挥了其抵抗炎症的作用,减弱了早期的放射性肺损伤。因此,我们认为Treg抑制后导致的Th1型细胞因子IFN-γ的增多可能是放射性肺损伤缓解的重要原因。有研究结果表明,IL-17A具有促进肺纤维化的功能[19-20],且中和小鼠体内的IL-17A能修复博来霉素引起的肺炎,减弱肺纤维化,提高小鼠的存活率[21-23]。受照后肺组织中IL-17A水平升高,这说明IL-17A在辐射诱导的肺损伤中起到了促进炎症的作用,并且加入CD25抗体后抑制了IL-17A的分泌,减缓了炎症的进程。Thl和Th2型细胞因子在其他因素引起的纤维化中发挥了重要作用[24-25],Th1型细胞因子(如IL-2和IFN-γ)可抑制纤维细胞分化[23, 26],而Th2型细胞因子(如IL-4和IL-10)可促进纤维细胞分化,并通过促进成纤维细胞的增殖和胶原蛋白的产生来调节纤维化组织修复[27]。在许多炎性病变的晚期,包括哮喘、硬皮病(全身性硬化)、心脏纤维化、肝纤维化、早期类风湿性关节炎、热损伤、肥厚性瘢痕和各种肺纤维化中,均发现了一种或多种促纤维化细胞因子(IL-4、IL-10、IL-13或TGF-β)水平的升高[28-30]。本研究结果显示IL-4水平在单纯照射后的第4周和第8周都是升高的,而在照射后4周,照射+CD25组中IL-4水平较照射+IgG组降低,这说明抑制Treg后,降低了IL-4水平,减缓了炎症的进程,但照射后8周,照射+CD25组中IL-4水平显著升高,这说明抑制Treg已经无法抑制纤维化的进程,这从小鼠皮肤损伤的表现中也可以得到证明。
Treg的分化对放射性肺损伤的影响不仅仅是通过CD25+Foxp3+Treg和CD25+NRP1+Treg的作用,Treg、Th17细胞和Th1/Th2型细胞分泌的多种细胞因子也参与其中。本研究结果将为预防或缓解放射性肺损伤提供更多的实验依据。
利益冲突 本研究由署名作者按以下贡献声明独立开展,不涉及任何利益冲突。
作者贡献声明 王蕊负责实验的实施、论文初稿的撰写;魏威、董卓负责小鼠Treg抑制模型的建立;陈志远负责论文的修改与校正;衣峻萱负责部分数据的整理与分析;刘羽欣、谷雨璐负责流式细胞术的检测;赵刚负责小鼠的照射;金顺子负责实验的设计、论文的指导。
Treg分化对放射性肺损伤的影响
Effect of Tregs differentiation on radiation-induced lung injury
-
摘要:
目的 探讨调节性T细胞(Treg)的分化对放射性肺损伤的影响及其作用机制。 方法 建立Treg抑制小鼠模型,按随机数字表法将C57BL/6小鼠分成4组:空白对照组、单纯照射组、照射+免疫球蛋白G(IgG)组和照射+CD25组,每组12只,除空白对照组外其余3组小鼠给予单次20 Gy X射线全胸照射,照射+IgG组和照射+CD25组小鼠每周腹腔注射IgG抗体和CD25抗体。分别于照射后第4周和第8周各处死小鼠6只,采用流式细胞术检测小鼠肺组织内CD25+Foxp3+Treg(Foxp3:叉头样转录因子3)的百分比以鉴定模型是否建立成功;采用Western blot法检测单纯照射组小鼠肺组织内神经纤毛蛋白1(NRP1)的表达;采用免疫荧光法检测每组小鼠肺组织内CD25+NRP1+Treg的百分比;拍照并观察每组小鼠皮肤的损伤情况,采用苏木精-伊红染色法检测小鼠肺组织的病理学改变;采用酶联免疫吸附测定法检测每组小鼠肺组织内转化生长因子β1(TGF-β1)、白细胞介素(IL)-17A、干扰素γ(IFN-γ)、IL-2和IL-4的水平变化。两组间比较采用独立样本t检验。 结果 流式细胞术检测结果显示,照射后第4周和第8周,单纯照射组小鼠肺组织内CD25+Foxp3+Treg百分比[(1.73±0.04)%、(2.13±0.15)%]均较空白对照组[(1.14±0.02)%、(1.70±0.06)%] 明显升高,差异均有统计学意义(t=−26.680、−4.545,P=0.000、0.010),抑制Treg后,第4周和第8周时照射+CD25组小鼠肺组织内CD25+Foxp3+Treg百分比[(0.72±0.14)%、(0.27±0.02)%]均较单纯照射组明显降低,差异均有统计学意义(t=5.296、37.538,均P=0.000)。Western blot结果显示,照射后第4周和第8周,单纯照射组小鼠肺组织内NRP1蛋白表达水平均较空白对照组升高,差异均有统计学意义(t=−7.341、−9.127,均P=0.000)。免疫荧光法检测结果显示,照射后第4周和第8周,单纯照射组小鼠肺组织内CD25+NRP1+Treg的百分比均较空白对照组升高,而照射+CD25组CD25+NRP1+Treg百分比均较单纯照射组降低,且差异均有统计学意义(t=8.926、14.457,P=0.001、0.000)。观察小鼠皮肤损伤程度后发现,照射后第4周和第8周,单纯照射组小鼠皮肤损伤严重,而照射+CD25组小鼠照射后第4周时皮肤基本完好,第8周时出现脱毛脱皮。病理学结果显示,照射后第4周和第8周,与空白对照组相比,单纯照射组小鼠的肺组织结构破坏,肺泡壁增厚,细胞外基质增多,而照射+CD25组小鼠的肺组织结构完整,肺泡壁纤细。酶联免疫吸附测定结果显示,与空白对照组相比,照射后第4周,单纯照射组小鼠肺组织内IL-17A和IL-4的水平均升高,差异均有统计学意义(t=−8.492、−15.796,P=0.001、0.000),照射后第8周,TGF-β1和IL-17A水平升高,差异均有统计学意义(t=−11.072、−7.167,P=0.000、0.002),IL-2水平在第4周和第8周时均降低,IFN-γ水平在第4周时升高,差异有统计学意义(t=−27.393,P=0.000),第8周时下降;与单纯照射组相比,照射+CD25组小鼠TGF-β1和IL-17A水平在第4周和第8周时均降低(t=6.037、4.524、5.496、4.772,均P=0.000),IFN-γ水平升高(t=−7.006、−12.565,P=0.002、0.000),差异均有统计学意义,而IL-2和IL-4水平在第4周时均降低,第8周时均明显升高,差异均有统计学意义(t=2.866、−9.090、8.833、−7.191,均P=0.000)。 结论 放射性肺损伤小鼠的肺组织中出现Treg分化,并增强分泌TGF-β1促炎因子,同时干扰辅助T细胞(Th1、Th2型)细胞因子的平衡来促进放射性肺损伤的发生。 -
关键词:
- T淋巴细胞,调节性 /
- T淋巴细胞,辅助诱导 /
- 模型,动物 /
- Th细胞平衡 /
- 放射性肺损伤
Abstract:Objective To investigate the effects of the differentiation of regulatory T cells (Tregs) on radiation-induced lung injury and its mechanism. Methods A mouse model that inhibits Tregs was established. C57BL/6 mice were divided into the control group, simple irradiation group, irradiation+IgG group, and irradiation+CD25 group according to the random number table method. Each group comprised 12 mice. The mice in the simple irradiation group, irradiation+IgG group, and irradiation+CD25 group were given a single 20 Gy X-ray full chest irradiation. The mice in the irradiation+IgG group and irradiation+CD25 group were intraperitoneally injected with IgG antibody and CD25 antibody every week, respectively. Six mice were killed at 4 and 8 weeks after irradiation respectively. Flow cytometry was used to detect the percentage of CD25+Foxp3+Tregs in the lung tissue of each group of mice to identify whether the model was established successfully. Then, Western blot was used to detect the expression of neuropilin 1(NRP1) in the lungs of the mice in the control group and irradiation group. Immunofluorescence was used to detect the percentage of CD25+NRP1+Tregs. Photos were taken to observe the skin damage of each group of mice. Hematoxylin eosin staining was used to detect the pathological changes of lung tissue. The secretion of cytokines, namely, transforming growth factor-beta 1 (TGF-β1), interleukin (IL)-17A, interferon-γ (IFN-γ), IL-2, and IL-4, in the lung tissue of each group was detected by enzyme-linked immunosorbent assay. Independent sample t-test was used for comparison between the two groups. Results The results of flow cytometry showed that the percentage of CD25+Foxp3+Tregs ((1.73±0.04)%, (2.13±0.15)%) in the lung tissue of mice in the simple irradiation group was significantly higher than that in the control group ((1.14±0.02)%, (1.70±0.06)%) (t=−26.680, −4.545; P=0.000, 0.010) at 4 and 8 weeks. After the inhibition of Treg cells, the percentage of CD25+Foxp3+Tregs ((0.72±0.14)%, (0.27±0.02)%) in the lung tissue of mice in the irradiation+CD25 group was significantly lower than that in the irradiation group alone (t=5.296, 37.538; both P=0.000). The result of Western blot showed that the expression of NRP1 protein in the lung tissue of mice in the irradiation group increased significantly (t=−7.341, −9.127; both P=0.000). The results of immunofluorescence showed that the proportion of CD25+NRP1+Tregs in the lung tissue of mice in the irradiation group was higher than that in the control group at 4 and 8 weeks after irradiation. The proportion of CD25+NRP1+Tregs in the irradiation+CD25 group was lower than that in the irradiation group (t=8.926, 14.457; P=0.001, 0.000). The skin lesions observed were severe in the simple irradiation group. The skin in the irradiated+CD25 group was almost intact at 4 weeks, and hair removal and peeling still occurred at 8 weeks. Hematoxylin eosin staining showed that relative to that in the control group, the lung tissue structure of the mice in the irradiated group was destroyed, the alveolar wall was thickened, and the extracellular matrix was increased at 4 and 8 weeks after irradiation. The lung tissue of the irradiated+CD25 group was intact and the alveolar wall was slender. As indicated by the changes in inflammatory factors, relative to that in the control group, the secretion of IL-17A and IL-4 in the lung tissue of mice increased in the simple irradiation group (t=−8.492, −15.796; P=0.001, 0.000) at 4 weeks. After 8 weeks of irradiation, the levels of TGF-β1 and IL-17A increased significantly (t=−11.072, −7.167; P=0.000, 0.002), and the levels of IL-2 decreased at 4 and 8 weeks. IFN-γ secretion increased at 4 weeks (t=−27.393, P=0.000) and decreased at 8 weeks. Relative to that in the simple irradiation group, TGF-β1 and IL-17A decreased (t=6.037, 4.524, 5.496, 4.772; all P=0.000), IFN-γ increased (t=−7.006, −12.565; P=0.002, 0.000) at 4 and 8 weeks, and IL-2 and IL-4 decreased at 4 weeks and increased significantly at 8 weeks (t=2.866, −9.090, 8.833, −7.191; all P=0.000) in the irradiation+CD25 group. Conclusion The differentiation of Tregs occurs in the lung tissue of mice with radiation-induced lung injury and promotes the development of radiation-induced lung injury by secreting TGF-β1 pro-inflammatory factor while interfering with the balance of helper T cell (Th)1/Th2 type cytokines. -
图 1 20 Gy X射线照射后C57BL/6小鼠Treg抑制模型肺组织中CD25+Foxp3+Treg的百分比
Figure 1. Percentage of CD25+Foxp3+Tregs in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
图 2 20 Gy X 射线照射后C57BL/6小鼠肺组织内NRP1蛋白的表达量变化
Figure 2. Changes of NRP1 protein in lung tissue of the C57BL/6 mouse irradiated by 20 Gy X-ray
图 3 20 Gy X 射线照射后C57BL/6小鼠Treg抑制模型肺组织内CD25+NRP1+Treg的百分比
Figure 3. Percentage of CD25+NRP1+Tregs in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
图 4 20 Gy X 射线照射后C57BL/6小鼠Treg抑制模型中各组小鼠的皮肤损伤及肺组织的病理学改变 图中,A:皮肤损伤的变化;B:肺组织的病理学变化(苏木精-伊红染色,×20)。IgG:免疫球蛋白G;Treg:调节性T细胞
Figure 4. Skin damage and pathological changes in lung tissue of the mouse Tregs inhibition model irradiated by 20 Gy X-ray
-
[1] Shi SM, Zeng ZC, Ye LX, et al. Risk Factors Associated With Symptomatic Radiation Pneumonitis After Stereotactic Body Radiation Therapy for Stage I Non-Small Cell Lung Cancer[J]. Technol Cancer Res Treat, 2017, 16(3): 316−320. DOI: 10.1177/1533034616661665. [2] Jin H, Kang GY, Jeon S, et al. Identification of molecular signatures involved in radiation-induced lung fibrosis[J]. J Mol Med (Berl), 2019, 97(1): 37−47. DOI: 10.1007/s00109−018−1715−9. [3] Veiga-Parga T, Sehrawat S, Rouse BT. Role of regulatory T cells during virus infection[J]. Immunol Rev, 2013, 255(1): 182−196. DOI: 10.1111/imr.12085. [4] Lo Re S, Lison D, Huaux F. CD4+ T lymphocytes in lung-x fibrosis: diverse subsets, diverse functions[J]. J Leukoc Biol, 2013, 93(4): 499−510. DOI: 10.1189/jlb.0512261. [5] Charbonnier LM, Chatila TA. Phenotypic and Functional Characterization of Regulatory T Cell Populations[M]//Soboloff J, Kappes DJ. Signaling Mechanisms Regulating T Cell Diversity and Function. Boca Raton: CRC Press, 2018: 105–118. [6] Fontenot JD, Rasmussen JP, Williams LM, et al. Regulatory T Cell Lineage Specification by the Forkhead Transcription Factor Foxp3[J]. Immunity, 2005, 22(3): 329−341. DOI: 10.1016/j.immuni.2005.01.016. [7] Gotot J, Dhana E, Yagita H, et al. Antigen-specific Helios-, Neuropilin-1-Tregs induce apoptosis of autoreactive B cells via PD-L1[J]. Immunol Cell Biol, 2018, 96(8): 852−862. DOI: 10.1111/imcb.12053. [8] Campos-Mora M, Morales RA, Pérez F, et al. Neuropilin-1+ regulatory T cells promote skin allograft survival and modulate effector CD4+ T cells phenotypic signature[J]. Immunol Cell Biol, 2015, 93(2): 113−119. DOI: 10.1038/icb.2014.77. [9] Clever D, Roychoudhuri R, Constantinides MG, et al. Oxygen Sensing by T Cells Establishes an Immunologically Tolerant Metastatic Niche[J]. Cell, 2016, 166(5): 1117−1131.E14. DOI: 10.1016/j.cell.2016.07.032. [10] Kamio K, Azuma A, Matsuda K, et al. Resolution of bleomycin-induced murine pulmonary fibrosis via a splenic lymphocyte subpopulation[J]. Respir Res, 2018, 19(1): 71. DOI: 10.1186/s12931−018−0783−2. [11] Gao YL, Chai YF, Qi AL, et al. Neuropilin-1highCD4+CD25+ Regulatory T Cells Exhibit Primary Negative Immunoregulation in Sepsis[J]. Mediators Inflamm, 2016, 2016: 7132158. DOI: 10.1155/2016/7132158. [12] Zheng SG, Wang JH, Horwitz DA. Cutting edge: Foxp3+CD4+CD25+ Regulatory T Cells Induced by IL-2 and TGF-β are Resistant to Th17 Conversion by IL-6[J]. J Immunol, 2008, 180(11): 7112−7116. DOI: 10.4049/jimmunol.180.11.7112. [13] Kong N, Lan Q, Su WG, et al. Induced T regulatory cells suppress osteoclastogenesis and bone erosion in collagen-induced arthritis better than natural T regulatory cells[J]. Ann Rheum Dis, 2012, 71(9): 1567−1572. DOI: 10.1136/annrheumdis−2011−201052. [14] Shao HY, Huang JY, Lin YW, et al. Depletion of regulatory T-cells leads to moderate B-cell antigenicity in respiratory syncytial virus infection[J]. Int J Infect Dis, 2015, 41: 56−64. DOI: 10.1016/j.ijid.2015.10.026. [15] Vignali D, Gürth CM, Pellegrini S, et al. IL-7 Mediated Homeostatic Expansion of Human CD4+CD25+FOXP3+ Regulatory T Cells After Depletion With Anti-CD25 Monoclonal Antibody[J]. Transplantation, 2016, 100(9): 1853−1861. DOI: 10.1097/TP.0000000000001276. [16] Wang LL, Wang XC, Tong L, et al. Recovery from acute lung injury can be regulated via modulation of regulatory T cells and Th17 cells[J]. Scand J Immunol, 2018, 88(5): e12715. DOI: 10.1111/sji.12715. [17] Wirsdörfer F, Cappuccini F, Niazman M, et al. Thorax irradiation triggers a local and systemic accumulation of immunosuppressive CD4+ FoxP3+ regulatory T cells[J/OL]. Radiat Oncol, 2014, 9: 98[2019-03-12]. https://ro-journal.biomedcentral.com/articles/10.1186/1748-717X-9-98. DOI: 10.1186/1748-717X-9-98. [18] Xu L, Xiong SS, Guo RF, et al. Transforming growth factor β3 attenuates the development of radiation-induced pulmonary fibrosis in mice by decreasing fibrocyte recruitment and regulating IFN-γ/IL-4 balance[J]. Immunol Lett, 2014, 162(1): 27−33. DOI: 10.1016/j.imlet.2014.06.010. [19] Wang A, Wang Z, Cao Y, et al. CCL2/CCR2-Dependent Recruitment of Th17 Cells but Not Tc17 Cells to the Lung in a Murine Asthma Model[J]. Int Arch Allergy Immunol, 2015, 166(1): 52−62. DOI: 10.1159/000371764. [20] Bui TT, Piao CH, Song CH, et al. Piper nigrum extract ameliorated allergic inflammation through inhibiting Th2/Th17 responses and mast cells activation[J]. Cell Immunol, 2017, 322: 64−73. DOI: 10.1016/j.cellimm.2017.10.005. [21] Chakraborty K, Chatterjee S, Bhattacharyya A. Impact of Treg on other T cell subsets in progression of fibrosis in experimental lung fibrosis[J]. Tissue Cell, 2018, 53: 87−92. DOI: 10.1016/j.tice.2018.06.003. [22] Cheng S, Chen HL, Wang AL, et al. Blockade of IL-23 ameliorates allergic lung inflammation via decreasing the infiltration of Tc17 cells[J]. Arch Med Sci, 2016, 12(6): 1362−1369. DOI: 10.5114/aoms.2016.62923. [23] Chen Y, Li C, Lu YP, et al. IL-10-Producing CD1dhiCD5+ Regulatory B Cells May Play a Critical Role in Modulating Immune Homeostasis in Silicosis Patients[J/OL]. Front Immunol, 2017, 8: 110[2019-03-12]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5303715. DOI: 10.3389/fimmu.2017.00110. [24] Sharma R, Sung SSJ, Gaskin F, et al. A novel function of IL-2: Chemokine/chemoattractant/retention receptor genes induction in Th subsets for skin and lung inflammation[J]. J Autoimmun, 2012, 38(4): 322−331. DOI: 10.1016/j.jaut.2012.02.001. [25] Liang P, Peng S, Zhang M, et al. Huai Qi Huang corrects the balance of Th1/Th2 and Treg/Th17 in an ovalbumin-induced asthma mouse model[J]. Biosci Rep, 2017, 37(6): BSR20171071. DOI: 10.1042/BSR20171071. [26] Xiong SS, Guo RF, Yang ZH, et al. Treg depletion attenuates irradiation-induced pulmonary fibrosis by reducing fibrocyte accumulation, inducing Th17 response, and shifting IFN-γ, IL-12/IL-4, IL-5 balance[J]. Immunobiology, 2015, 220(11): 1284−1291. DOI: 10.1016/j.imbio.2015.07.001. [27] Grabarz F, Aguiar CF, Correa-Costa M, et al. Protective role of NKT cells and macrophage M2-driven phenotype in bleomycin-induced pulmonary fibrosis[J]. Inflammopharmacology, 2018, 26(2): 491−504. DOI: 10.1007/s10787−017−0383−7. [28] Sziksz E, Pap D, Lippai R, et al. Fibrosis Related Inflammatory Mediators: Role of the IL-10 Cytokine Family[J]. Mediators Inflamm, 2015, 2015: 764641. DOI: 10.1155/2015/764641. [29] Skuljec J, Jirmo AC, Habener A, et al. Absence of Regulatory T Cells Causes Phenotypic and Functional Switch in Murine Peritoneal Macrophages[J/OL]. Front Immunol, 2018, 9: 2458[2019-03-12]. https://www.frontiersin.org/articles/10.3389/fimmu.2018.02458/full. DOI: 10.3389/fimmu.2018.02458. [30] Villanueva-Romero R, Gutiérrez-Cañas I, Carrión M, et al. The Anti-Inflammatory Mediator, Vasoactive Intestinal Peptide, Modulates the Differentiation and Function of Th Subsets in Rheumatoid Arthritis[J]. J Immunol Res, 2018, 2018: 6043710. DOI: 10.1155/2018/6043710. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3356
- HTML全文浏览量: 2329
- PDF下载量: 14
